Article Text

Abstract
Objective and method: To outline the direct and indirect approaches in the fight against blood doping in sports, the different strategies that have been used and are currently being used to fight efficiently against blood doping are presented and discussed.
Results and conclusions: The paper outlines the different approaches and diagnostic tools that some federations have to identify and target sportspeople demonstrating abnormal blood profiles. Originally blood tests were introduced for medical reasons and for limiting misuse of recombinant human erythropoietin (rHuEPO). In this way it became possible to prevent athletes with haematocrit levels well above normal, and potentially dangerous for their health, competing in sport. Today, with nearly a decade of blood testing experience, sports authorities should be familiar with some of the limitations and specially the ability of blood tests performed prior to competitions to fight efficiently against the misuse of rHuEPO, blood transfusion, and artificial haemoglobin.
- BFU-E, burst forming unit-erythroid
- CFU-E, colony forming unit-erythroid
- EPO, erythropoietin
- IOC, International Olympic Committee
- LAD, Swiss Laboratory for Doping Analyses
- rHuEPO, recombinant human EPO
- sTFR, soluble transferring receptor
- EPO
- doping
- transfusion
- haematocrit
- reticulocyte count
Statistics from Altmetric.com
- BFU-E, burst forming unit-erythroid
- CFU-E, colony forming unit-erythroid
- EPO, erythropoietin
- IOC, International Olympic Committee
- LAD, Swiss Laboratory for Doping Analyses
- rHuEPO, recombinant human EPO
- sTFR, soluble transferring receptor
Erythropoiesis is part of the large process of haematopoiesis, which involves the production of mature cells found in the blood and lymphoid organs.1 Haematopoiesis is continuously required because of the normal turnover in the cell populations in the blood and lymphoid organs. In the normal adult human, the daily turnover of erythrocytes exceeds 1011 cells. During periods of increased erythrocyte loss, due to haemolysis or haemorrhage, the production of erythrocytes increases rapidly and markedly. However, overproduction of erythrocytes does not occur, even after the most severe loss of erythrocytes.
In haematopoiesis, a few pluripotent haematopoietic stem cells in the bone marrow proliferate and differentiate to give rise to all the cellular components of the blood and the lymphoid system. During this process, an individual haematopoietic cell undergoes an apparently random process called commitment. When a cell undergoes commitment, its potential to proliferate becomes limited and its potential to develop into multiple types of mature cell is also restricted. Thus, these haematopoietic cells are termed committed, lineage specific progenitor cells.
The major stages of differentiation in mammalian erythropoiesis are as follows. The most immature stage of committed erythroid progenitors is the burst forming unit-erythroid (BFU-E). The next major stage of erythroid progenitor cell development is the colony forming unit-erythroid (CFU-E). A continuum of erythroid progenitor stages exists between the BFU-E and CFU-E, with decreasing proliferative potential as the progenitors approach the CFU-E stage. The descendant cells of the CFU-E are termed erythroid precursor cells. These erythroid precursors are proerythroblasts, basophilic erythroblasts, polychromatophilic erythroblasts, and orthochromatic erythroblasts. The orthochromatic erythroblasts do not divide, but they enucleate, forming the nascent erythrocyte called the reticulocyte.
PRODUCTION OF ERYTHROPOIETIN
Erythropoietin (EPO) is a 30 400 molecular weight glycoprotein hormone produced mainly in the kidney, and also in the liver (<10%) and, in very little quantities, in the brain.2–5 The physiological stimulus for EPO production is tissue hypoxia, which, in the large majority of instances, is directly related to the number of circulating erythrocytes.6 Thus, EPO and erythropoiesis are part of a negative feedback cycle that keeps tissue oxygen delivery within a narrow range by controlling the number of erythrocytes circulating in the blood.7 ln a normal individual, any loss of erythrocytes, such as by bleeding or haemolysis, decreases delivery of oxygen to the tissues.8 When this tissue hypoxia is sensed by cells in the kidney and liver capable of producing EPO, they produce and secrete EPO into the plasma.9 EPO is carried to the bone marrow, where it binds to specific cell surface receptors on its target cells—the CFU-E, pro-erythroblasts, and basophilic erythroblasts.10,11 The binding of EPO by these cells increases their ability to survive and reach the reticulocyte stage and thereby contribute to the population of circulating erythrocytes. The increased numbers of circulating erythrocytes in turn deliver more oxygen to the tissues. This increased oxygen delivery is sensed by the EPO producing cells, which then reduce EPO production so that the normal steady state number of erythrocytes is restored.
The response of the kidneys to hypoxia has been shown to be exponential12—that is, in individuals with a normal capacity to produce EPO, a linear decline in haematocrit is accompanied by an exponential increase in plasma EPO levels. This exponential increase is not based on the release of stored, preformed EPO. Rather, the hypoxia is sensed by an intracellular molecule that interacts with an enhancer element of the Epo gene and thereby induces transcription of the gene.13 The increase in EPO production in the hypoxic kidney is achieved by recruitment of more cells to produce EPO. The EPO producing cells of the kidney are a minor subset of cortical interstitial cells. Under normal conditions, only a few scattered cells produce EPO. When a threshold level of hypoxia is achieved, the cells capable of producing EPO do so at a maximal rate. The greater the areas of renal cortex in which the hypoxia threshold is met, the greater the number of cells that produce EPO.9
MECHANISM OF ACTION OF ERYTHROPOIETIN
ln the bone marrow, EPO binds to receptors displayed on the cell surface of CFU-E, proerythroblasts, and basophilic erythroblasts. The mature EPO receptor, with a molecular weight of approximately 72 000, is a transmembrane glycoprotein, a member of a much larger family of receptors of cytokines and haematopoietic growth factors. The effect of EPO binding to its receptor, in terms of cellular physiology, has been shown to be the prevention of programmed cell death (apoptosis).14,15 In multiple systems of erythropoiesis, EPO has been shown to be a survival factor for the erythroid cells in the later stages of differentiation from the CFU-E through basophilic erythroblasts. Although an effect of EPO on mitosis has been reported for BFU-E and an EPO dependent cell line, EPO is required only for CFU-E and later stages, and apoptosis appears to result when EPO signalling cannot occur.
DETECTING RHUEPO MISUSE IN SPORTS
The availability of recombinant human EPO (rHuEPO) in 1987 in Europe made it clear that this ergogenic hormone would be used illicitly in endurance sports. Therefore, the International Olympic Committee (IOC) Medical Commission decided to ban this drug in 1990, even though all forms of blood doping had been officially banned since 1984. Two philosophies were developed for the detection of rHuEPO misuse in sports. The first one was based on the detection of indirect blood markers and the second one was based on the direct detection of rHuEPO in urine.16 The promotion of secondary blood markers was mainly on the basis that they could be used to detect rHuEPO injected a long time ago (more than a week ago), and also that they could be used to detect all kinds of erythropoietic stimulator such as erythropoietin alfa, beta, omega, and delta, and darbepoetin alfa and mimetic peptides.17,18 Furthermore, secondary blood markers could eventually be used to identify athletes who ceased using rHuEPO or other erythropoietic stimulators. In the meantime, scientists were working on the direct detection of rHuEPO in blood or urine. This latter method had the advantage of identifying the drug itself (or metabolites), but had the disadvantage of being expensive, little sensitive, and delicate to perform.
Indirect methods of detection
In 1993, with the introduction of sophisticated haematological analysers some scientists proposed a model implicating the analysis of the percentage of red blood cells having a haemoglobin concentration below 28 pg (mean corpuscular haemoglobin (MCH)) and a volume above 128 fl (mean corpuscular volume (MCV)). These red blood cells were called macrocytic hypochromatic erythrocytes. This test had the advantage of being fast and cheap (as long as the laboratory was equipped with this special analyser) and was highly selective. Unfortunately, the test was limited by its relatively poor sensitivity; 50% of the rHuEPO samples were not detected.19
Another indirect test, developed in 1996 for the detection of rHuEPO misuse, was based on the determination of the soluble transferring receptor (sTFR)/ferritin ratio.20 The results of a trial involving healthy subjects demonstrated that regular rHuEPO injections significantly increased the sTFR concentration. Ferritin was used as a denominator mainly to prevent variations in hydration level. Unfortunately, during this trial, the ferritin levels of the subjects collapsed because they did not receive any iron supply. As iron supplementation is a common practice among athletes (specially intravenous iron injections),21 the sTFR/ferritin ratio was modified into a new ratio taking into account the possible exercise induced haemoconcentration, the sTFR/total protein.22
The lack of sensitivity of some of the secondary blood markers as well as the lack of specificity of others encouraged some scientists to put them together in a multiple markers mathematical model to discriminate rHuEPO misusers from healthy sportspeople. Following a double blind study with regular rHuEPO injections (continuous treatment), the Australian Institute of Sport, together with the Australian Sports Drug Testing Laboratory, designed an anti-doping test using multiple secondary blood markers such as the haematocrit level, the reticulocyte haematocrit, serum sTFR and EPO concentrations, and lastly the percentage macrocytic cells. Different mathematical models were developed allowing the identification of sportspeople under rHuEPO treatment (ON-model) and those who had taken rHuEPO in the past (OFF-model).23 In August 2000, the IOC Medical Commission approved the ON-model to be used during the 2000 Olympic Games in Sydney. As the direct method capable of discriminating endogenous EPO from rHuEPO had already been published in spring 2000,24 the ON-model was used only as a screening test to determine which urine samples had to be collected to perform the urinary test.
At the same time that the above mentioned study was performed in Australia, the Laboratoire Suisse d’Analyse du Dopage (Swiss Laboratory for Doping Analyses; LAD) conducted a similar randomised controlled, double blind trial, except that iron supplementation was given much more importance and it was given intravenously—for maximum efficiency and to be as close as much as was possible to what is done in cycling. The results showed that the behaviour of secondary blood markers was different during the continuous treatment. In contrast with the Australian study, the Swiss study demonstrated that some of the secondary blood markers (haematocrit, haemoglobin, and reticulocyte count) could be used as part of a screening test, but in no case could be used for anti-doping purposes (fig 1).25
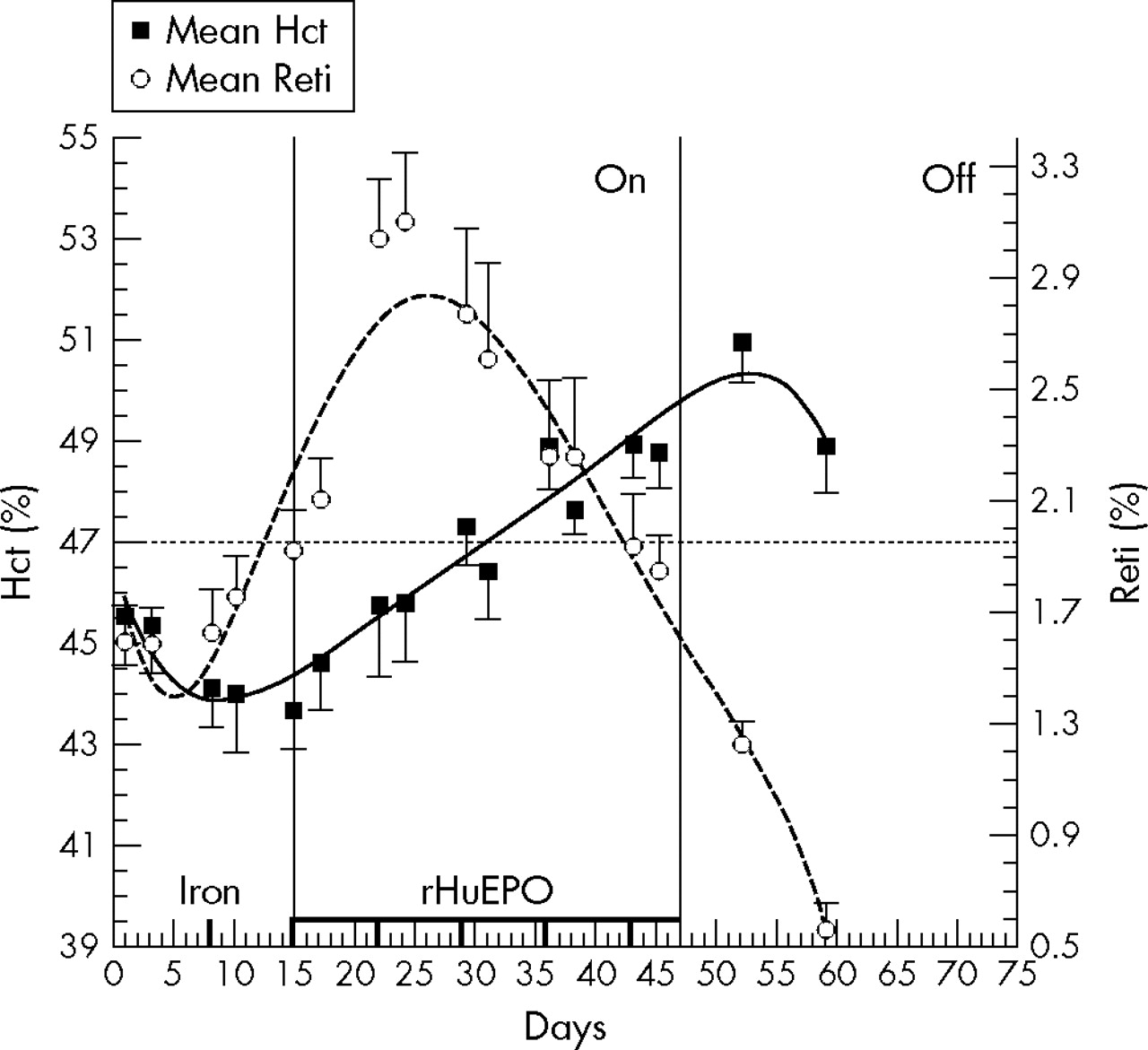
Model showing the profile of mean haematocrit (Hct) and reticulocyte count (Reti) (standard error of mean) during continuous recombinant human erythropoietin (rHuEPO) treatment with regular intravenous iron injections (see vertical bars near the x axis). The study included three periods: baseline, treatment (ON) and post-treatment (OFF). The horizontal line represents a theoretical cut-off limit to determine which urine samples should be collected to confirm rHuEPO misuse.
Direct methods of detection
Endogenous EPO and rHuEPO are slightly different, and these differences are certainly because the glycosylation of rHuEPO takes place in Chinese Hamster Ovary (CHO) cells rather than in human cells.26 Indeed, the post-translational modifications are species and tissue dependent and also dependent on the cell culture conditions. Therefore, it is possible to separate the endogenous from the exogenous EPO isoforms based on the differences in the charge status of different sugars.27,28 The technique developed by Wide allowed the separation of the various isoforms thanks to the differences in charge status of the different sugars. Is was shown that this technique was reliable in urine and in blood as long as the biological samples were collected within 24 hours after the last rHuEPO injection. Unfortunately, once the rHuEPO treatment had ceased for more than three days, less than 50% of the treated subjects could be declared positive. Seven days after the last rHuEPO injection, none of the samples showed any traces of rHuEPO.29,30
A few months before the 2000 Summer Olympic Games in Sydney, the French anti-doping laboratory in Paris published in Nature a novel test based on isoelectric focusing patterning and a double blotting protocol.24 As the exogenous isoforms of rHuEPO are less acidic than the endogenous EPO, it was possible to develop a convenient protocol to separate them using the isoelectric focusing method. This test was designed to separate alfa, beta, and omega rHuEPO as well as darbepoetin alfa (see fig 2).31,32
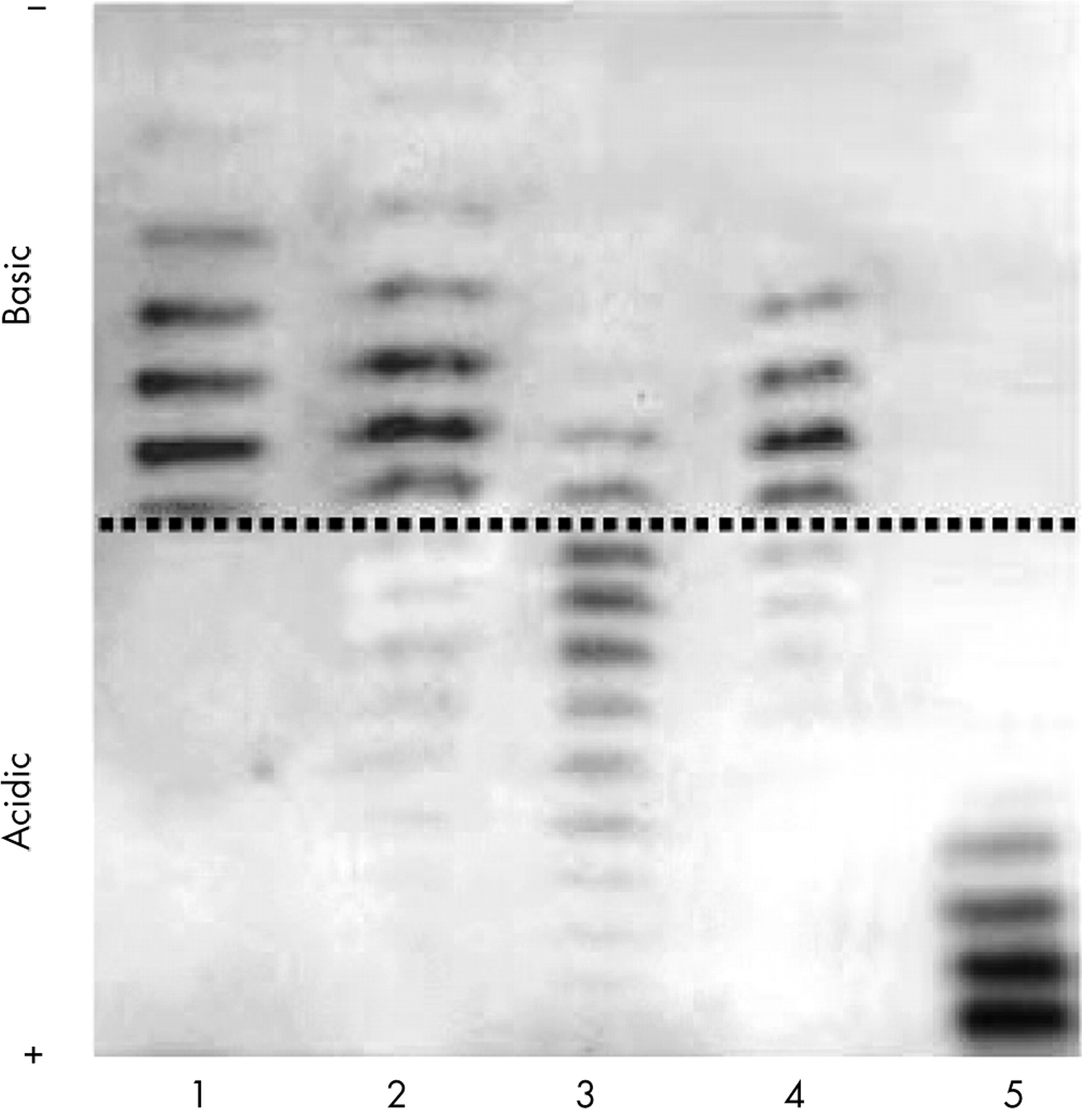
Anti-doping urine analysis demonstrating the presence of recombinant human erythropoietin (rHuEPO) in urine (see lane 4). Lane 1: rHuEPO standard; 2: positive urine (control); 3: negative urine (control); 4: sample declared positive; 5: darbepoetin alfa (Aranesp standard).
TARGETING RHUEPO MISUSERS
The LAD and the some federations took the decision together to launch the blood screening test based on the determination of the haematocrit, the haemoglobin and the reticulocyte count. It was introduced during the 2001 cycling season, Tour des Flandres.33 Very soon it was demonstrated that blood test was capable of identifying rHuEPO misusers.23,34–36 Since then, quite a few other sports federations have decided to introduce the screening test set up in Lausanne. With time, this test was shown to be even more efficient during a follow up of the athletes (blood profile), and variations above normal were shown to be excellent indicators of blood manipulation. In fact some models take these variations into account, and athletes demonstrating variations above their individual reference range are targeted and have to go through appropriate urine and blood anti-doping tests.37
What is already known about this topic
Blood doping has been common practice in some endurance sports for quite a few decades, because its efficiency has clearly been demonstrated. In the early 1990s, rHuEPO was launched on the market. As a side effect of this blood doping became even more attractive. For that reason, some international sports federations had to limit and fight against blood doping, notably by analysing blood markers of altered erythropoiesis, such as haematocrit, haemoglobin, and reticulocyte count. Finally in 2000, a direct detection test of rHuEPO in urine was introduced which enabled demonstration of the origin of EPO.
ABNORMAL BLOOD PROFILES
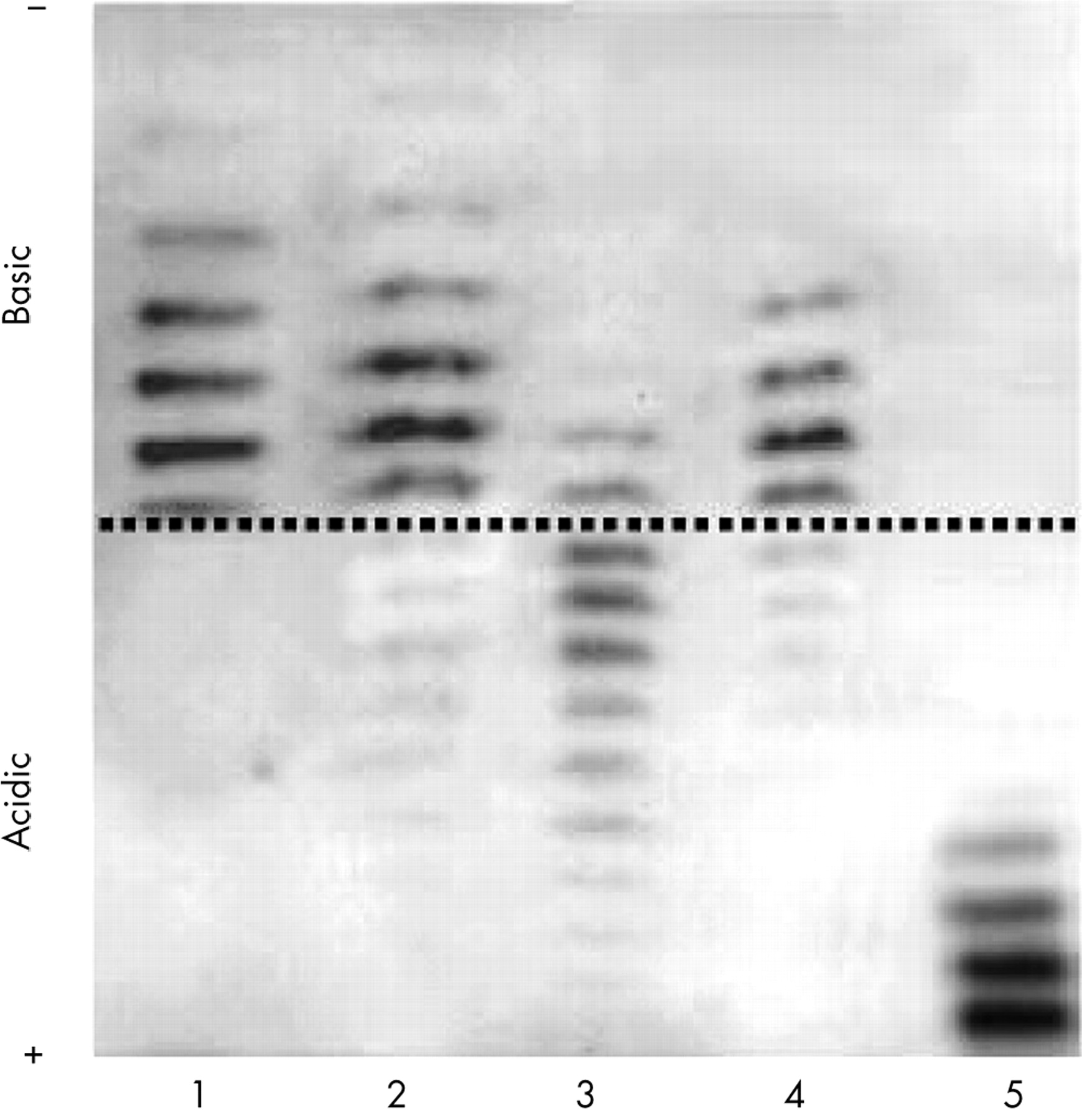
For approximately two years, abnormal blood profiles have been noticed without any traces of rHuEPO in urines.36 This meant that athletes were either doping with undetectable compounds or had returned to “old” doping techniques, such as blood transfusion. Blood transfusion to enhance oxygen transport with the increase of red blood cell mass was common practice in the early 1970s.38 This way of doping virtually disappeared with the arrival of rHuEPO on the market at the end of the 1980s, because it is much easier to use the hormone (easy to store and to use). The launching in 2000 of the direct detection test of rHuEPO in urine samples had a disadvantage—a return to blood transfusion. The regular follow up of blood markers such as haematocrit, haemoglobin, and reticulocyte count suddenly showed that some athletes had abnormal values of blood markers although rHuEPO could not be detected in urine. This clearly indicated a return to blood transfusion practices. With the possibility of analysing specifically the red blood cell membrane proteins defining notably the different blood groups and subgroups, the LAD decided to perform for the first time anti-doping tests in blood (summer 2004) (fig 3). The specific labelling of some red blood cell membrane proteins in combination with flow cytometry proved that abnormal blood profiles were due to homologous blood transfusion.39 Thus the disadvantage of anti-doping tests is that some sportspeople change their doping habits, notably they recourse to autologous blood transfusion. This method is for the time being undetectable, but the policy of “No Start” introduced by some federations will strongly limit the efficiency of this doping strategy (see reference 37).

{kind=link}
{kind=link}
{kind=link}
Anti-doping blood analysis demonstrating the presence of a single (A, B) or mixed (C, D) red blood cell population(s). A: single population without expression; B: single population with expression; C: mixed population with a majority expressing; D: mixed population with the majority non expressing. FITC, fluoroscein isothiocynate.
The appropriate way to fight blood doping is to establish for each marker and measure (haematocrit, haemoglobin, OFF-score, reticulocyte count) individual reference values.37 In this way it will not be necessary to take into account the sex, ethnic origin and the kind of sport practised. Each person has his or her own set reference values and based on variations and evolution of the markers over time, it is possible to identify those athletes who are manipulating. This is certainly the best way to fight blood doping as long as the preanalytical and analytical conditions are optimal. The approach improves even more when announced and unannounced blood data are put together, because data of athletes manipulating their blood at the time of announced blood controls will show discrepancies. Eventually, in the future, a policy of “No Start” for all athletes demonstrating abnormal blood values could be a solution to limit the costs related to the elaboration of specific anti-doping tests for all imminent molecules/methods and will nevertheless limit the misuse of blood doping.17,18,40
What this study adds
Since the introduction of rHuEPO on the market, quite a few strategies have been elaborated to fight blood doping. This review describes the strategies, their limitations, and their potential to fight blood doping more efficiently, notably homologous blood transfusion.
CONCLUSIONS
Sports federations that have introduced blood tests actually have a powerful tool to follow all athletes potentially misusing rHuEPO or blood transfusion. The anti-doping tests need to be focused mainly on those demonstrating abnormal blood data. This targeting will also enable the federations to determine the prevalence of doping methods before any validated anti-doping test is introduced on the market. This was the way in which it was found out that haemoglobin based oxygen carrier (HBOC) misuse was not a major problem.
REFERENCES
Footnotes
-
Competing interests: none declared