Article Text

Abstract
Background—Pulmonary diffusing capacity for carbon monoxide (Dlco), alveolar capillary membrane diffusing capacity (Dm), and pulmonary capillary blood volume (Vc) are all significantly reduced after exercise.
Objective—To investigate whether measurement position affects this impaired gas transfer.
Methods—Before and one, two, and four hours after incremental cycle ergometer exercise to fatigue, single breath Dlco, Dm, and Vc measurements were obtained in 10 healthy men in a randomly assigned supine and upright seated position.
Results—After exercise, Dlco, Dm, and Vc were significantly depressed compared with baseline in both positions. The supine position produced significantly higher values over time for Dlco (5.22 (0.13) v 4.66 (0.15) ml/min/mm Hg/l, p = 0.022) and Dm (6.78 (0.19) v 6.03 (0.19) ml/min/mm Hg/l, p = 0.016), but there was no significant position effect for Vc. There was a similar pattern of change over time for Dlco, Dm, and Vc in the two positions.
Conclusions—The change in Dlco after exercise appears to be primarily due to a decrease in Vc. Although the mechanism for the reduction in Vc cannot be determined from these data, passive relocation of blood to the periphery as the result of gravity can be discounted, suggesting that active vasoconstriction of the pulmonary vasculature and/or peripheral vasodilatation is occurring after exercise.
- pulmonary
- diffusing capacity
- exercise
- body position
- blood volume
- lung
Statistics from Altmetric.com
Take home message
The reduction in pulmonary diffusing capacity after exercise was not alleviated by altering body position, and was primarily due to a decrease in pulmonary capillary blood volume. The data indicate that the reduction in the latter was not due to passive relocation of blood, suggesting that active vasoconstriction of the pulmonary vasculature and/or peripheral vasodilatation may be occurring after exercise.
The effect of changing the body position from seated to supine is to increase the diffusing capacity for carbon monoxide of the lung (Dlco). This increase has traditionally been attributed to an increase in the pulmonary capillary blood volume (Vc) in the upper pulmonary lobes.1 However, the capillaries in the upper regions of the lung are not fully recruited because of the relatively low vascular pressure. Thus, although Vc is substantially elevated, the rise in the alveolar capillary membrane diffusing capacity (Dm) is insignificant.2
Dlco is impaired after exercise of different durations and intensities.3–5 The restriction of Dlco after exercise persists for at least six hours and may not return to baseline values for one to two days.6–8 It reflects a decrease in both Dm and Vc.5,6
Blood pooling in the periphery after exercise and therefore compensatory shunting, as transient changes in venous tone in both pulmonary and systemic circulations direct blood flow away from the thorax to clear the metabolic byproducts of exercise, have been suggested to explain the decrement in Vc.6,9,10 Thoracic electrical impedance is elevated after exercise, indicating a lower thoracic fluid volume8,11 and supporting the theory about reduced central blood volume.
Microvascular injury to the alveolar capillary membrane resulting in acute subclinical interstitial pulmonary oedema has been suggested to be a mechanism for the reduced Dm.3,12–14 Maximal hydrostatic pressures in human pulmonary capillaries are not accurately known, although they are estimated to be in the region of 30–40 mm Hg.15 These pressures are in the range where stress failure of the pulmonary capillaries has been demonstrated in experimental animals.12,16 When pressures this high develop, the vascular endothelium may be injured allowing the movement of fluid from the vascular space to the interstitium of the lung. Pulmonary oedema could also result from an increased blood volume in the distended lung capillaries increasing permeability and promoting fluid shifts.7 An associated inability of the pulmonary lymphatic system to clear the accumulated fluid may explain the depressed Dm after exercise .17
If a change in body position from seated to supine at rest results in increased venous return and thereby promotes an increase in blood in the pulmonary capillaries, then a supine body position after exercise should also promote a reduced Vc. Hanel et al,9 investigating the redistribution of blood from the central vascular bed to more distal regions after exercise, reported a decrease in Dlco in both seated and supine positions (6% and 12% respectively).9 A corresponding change in thoracic to thigh electrical impedance ratio (+14%), in the seated position only, supported the authors' hypothesis explaining the decrease in Dlco after exercise. The seated and supine measurements were conducted in the same testing session, separated by only 15 minutes, despite the recovery being solely in a supine position.
The independent changes in Dlco associated with a change in body position and recovery from exercise have been confirmed on many occasions,2,8,18 but the interaction of these two treatments remains unresolved and forms the purpose of this investigation. If the decrease in Dlco and Dm after exercise were a function of a relocation of blood into the periphery causing a decrease in Vc, it was hypothesised that placing the subject in a supine position would enhance venous return after exercise and limit the decrease in central blood volume. This would return Vc toward baseline supine levels compared with the seated levels which would remain depressed.
Methods
Ten healthy non-smoking men were recruited. All were required to have normal pulmonary function with no known history of cardiovascular or respiratory disease. Before any testing, subjects received a verbal description of the experiment and completed a written consent form. This study was approved by the clinical screening committee for research and other studies involving human subjects of the University of British Columbia.
EXPERIMENTAL PROTOCOL
Subjects were required to report to the laboratory on two separate occasions, separated by at least 72 hours. Each testing session followed a similar protocol. To stabilise the pulmonary system, subjects were required to rest in their assigned position for 15 minutes before the measurement of Dlco.19 Pulmonary function was assessed by a Collins survey tach pulmonary function testing unit (Collins DS/PLUS II; Warren E Collins Inc, Baintree, Massachusetts, USA). Baseline values of Dlco were also obtained. A maximal cycle ergometer test was then undertaken. A total of five diffusion measurements were made, including one baseline measure and four measurements at 15 minutes, one, two, and four hours after the maximal cycle test. The two testing sessions differed only in the body position of the subject during the diffusion and pulmonary function measurements. Two positions were investigated, supine and seated. At all diffusion measurement periods, (a) heart rate was recorded using a telemetric monitor (Polar Vantage XL; Polar Electro Oy, Kempele, Finland) to evaluate a resting cardiovascular state, and (b) haemoglobin was measured using a direct reading haemoglobinometer (HemoCue, Helsingborg, Sweden) to correct Dlco measures.17
MAXIMAL CYCLE ERGOMETER TEST
For the maximal cycle exercise test, subjects cycled on an electronically braked cycle ergometer (Quinton Excalibur; Lode, Groningen, The Netherlands) at a self selected cadence (about 30–120 rpm), against a workload increasing at 30 W/min until volitional fatigue. Expired gases were collected through a two way non-rebreathing valve (Hans-Rudolph, no 2700B, Kansas City, Missouri, USA) and analysed with an automated metabolic system (Rayfield System consisting of an airflow meter (Vacumetrics no 17150, Ventura, California, USA), and S-3A oxygen analyser and a carbon dioxide analyser (Ametek, Pittsburg, Pennsylvania, USA)). Before each testing session, the gas analysers were calibrated with a known gas mixture and room air. Heart rate was recorded every 15 seconds, and arterial oxygen saturation (Sao2) was measured with a pulse ear oximeter (Ohmeda Biox 3740; BOC Health Care Inc, Edison, New Jersey, USA).
COLLECTION OF PULMONARY DIFFUSION DATA
Pulmonary diffusing capacity was determined by the single breath method,20,21 which required the subjects to make a maximal inspiration from residual volume of a gas mixture containing 20.9% O2, 9.7% He, 0.3% CO balanced with N2. The breath was held for about 10 seconds, and then expired. The first litre of expired gas was discarded, and the next 750 ml was considered to be an alveolar sample uncontaminated by dead space gas. Concentrations of CO were measured using an infrared analyser (type 101; Morgan, Kent, UK). Each diffusion measurement was examined to ensure that the inspired volume was at least 90% of the forced vital capacity, the total time of inspiration was less than two seconds, and breath hold time was between 9 and 11 seconds.21 Measurements were made in duplicate separated by five minutes to ensure elimination of the test gas from the lungs. Both tests were within 10% or 3 ml/min/mm Hg or a third test was performed.22 The average of the two closest values was recorded. The diffusion apparatus was calibrated daily for both volume and CO.
To calculate Dm and Vc, a second Dlco (Dlco 90% O2) test was performed. Subjects were required to breathe for five minutes through a low resistance valve (Hans Rudolph, no 2700B) attached to a Douglas bag filled with a gas mixture of about 90% O2 and 10% N2. The Dlco 90% O2 test was immediately performed in the same manner as the 21% O2. The 10 second breath hold was of a gas mixture of 90% O2, 10% He, and 0.3% CO. Dm and Vc were then determined.20 The partitioning technique is consistent with that performed by numerous authors,5,6 and the reliability has been shown by test-retest correlation for both gas measures (21% and 90% O2) to be r = 0.98 and r = 0.96 respectively.23
STATISTICAL ANALYSIS
Data were examined using a 2 (position) by 5 (time) factorial analysis of variance with repeated measures on both factors. Time effects were analysed using the Dunnet test for multiple comparisons to a control group, in which means after exercise were compared with resting values. If a significant interaction occurred, Scheffe's post hoc procedure was applied for further comparison. Student's t tests were used to compare data for resting pulmonary function and other descriptive data in both positions. The level of significance was p<0.05 for all statistical comparisons.
Results
Ten healthy men completed the study (mean (SD) age 22.3 (2.4) years, height 180.3 (8.9) cm, and weight 80.4 (8.6) kg). Data on resting respiratory function showed no signs of abnormality (table 1).
Resting pulmonary function data for subjects in the two measurement positions
MAXIMAL CYCLE ERGOMETER TESTS
Data for the two incremental ergometer tests to volitional fatigue showed that equal stress was applied to the subjects at each testing session (table 2). No subjects in this study were classified as having exercise induced hypoxaemia (an arterial oxygen saturation of less than 91%).24
Metabolic variables during maximal cycle ergometer test
CARDIOVASCULAR VARIABLES
The significantly elevated heart rate (46%) obtained immediately before the 15 minute Dlco determination indicated that, regardless of body position, the metabolic level in some subjects had not returned to that found before exercise (table 3). The 15 minute data for Dlco, Dm, and Vc were therefore removed from the analyses, because a resting cardiovascular state was required for standardised comparisons. This resulted in a position effect becoming significant for Dlco (p = 0.022 v p = 0.058 for the full analysis). There was no other significant change in Dlco, Dm, or Vc, when compared with the full analysis.
Heart rate (beats/min) at rest (baseline) and 15 minutes and one, two, and four hours after maximal exercise measured in the seated or supine position
PULMONARY DIFFUSION MEASUREMENTS
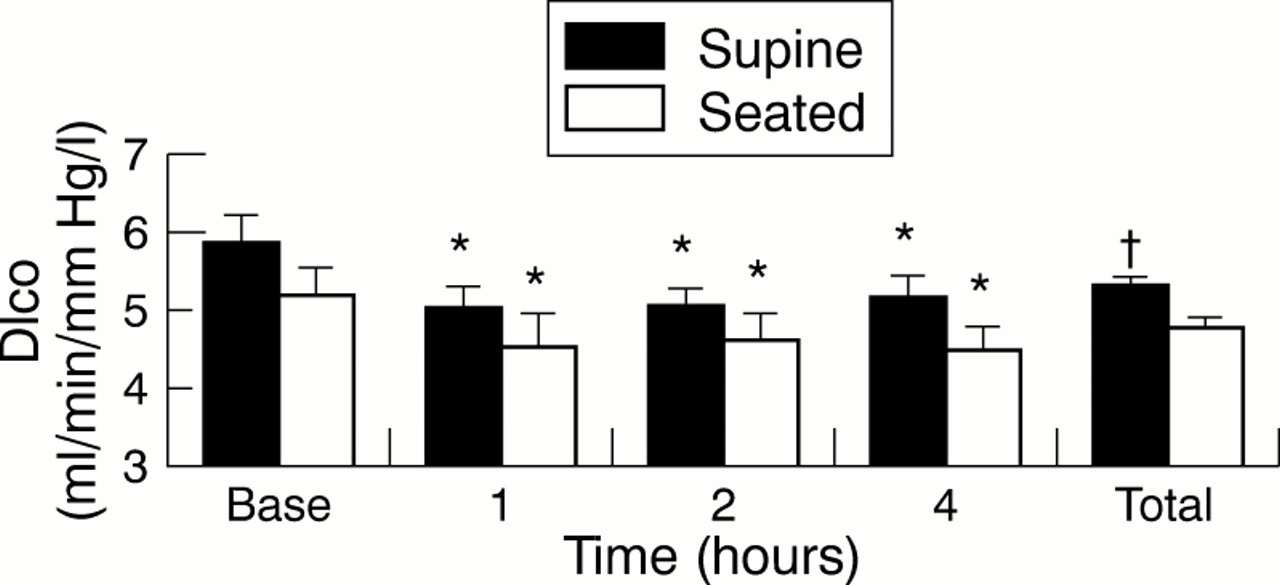
Dlco was significantly higher in the supine than the seated measurements (5.22 (0.13) v 4.66 (0.15) ml/min/mm Hg/l; p = 0.022). Both positions showed significant reductions in Dlco at one, two, and four hours after exercise (fig 1).

Pulmonary diffusing capacity for carbon monoxide (Dlco) corrected for alveolar volume at rest (Base), after exercise, and for each position regardless of time (Total). Values are means (SE). *Significantly lower than baseline value for the same position; †significantly higher than seated value; p<0.05.
Vc and Dm were both significantly lower than baseline at one, two, and four hours after exercise (figs 2 and 3 respectively). Dm showed a body position effect, with the supine position producing higher values (6.78 (0.19) v 6.03 (0.19) ml/min/mm Hg/l, p = 0.016). Vc was not significantly different between positions (13.49 (0.65) v 12.88 (0.60) ml/l, supine v seated; p = 0.623). The two positions exhibited a similar pattern of change over the time periods for Dlco, Dm, and Vc.

Pulmonary capillary blood volume (Vc) corrected for alveolar volume at rest (Base), after exercise, and for each position regardless of time (Total). Values are means (SE). *Significantly lower than baseline value for the same position; p<0.05.

{kind=link}
{kind=link}
{kind=link}
Membrane diffusing capacity (Dm) corrected for alveolar volume at rest (Base), after exercise, and for each position regardless of time (Total). Values are means (SE). *Significantly lower than for baseline value for the same position; †significantly higher than seated value; p<0.05.
Discussion
This study confirmed that pulmonary diffusing capacity is impaired during recovery from maximal exercise and that this reduction persists for at least four hours (fig 1).6,11 It also supports the recent finding that the impairment occurs in both a supine and upright seated position.9
PULMONARY CAPILLARY BLOOD VOLUME
Maximal exercise resulted in a reduced Vc in both a supine and upright seated position (fig 2). Position had no effect on the magnitude of the decrement over time, which was maintained four hours after exercise. The reduced Vc has been reported after exercise of various intensities, durations, and modalities.3–5,11 It has also been documented to remain depressed for a period of 24 hours.6,7
It has been suggested that a reduced central blood volume is the primary mechanism for the impaired Dlco after exercise.9,11 The decreased Vc found in our study (fig 2) supports this theory. Thoracic fluid volume, measured by electrical impedance, is decreased after exercise and has been shown to be still decreased two to three hours after exercise.8,9,11 A parallel relation between the decrease in Vc and an increase in thoracic electrical impedance indicates a reduction in central fluid volume, specifically blood. However, the mechanism behind this reduction in central blood volume remains unknown.
Inactivity for a prolonged period removes the effect of the muscle pump that assists venous return, suggesting that peripheral pooling of blood may be the cause of the decreased Vc after exercise. Even without a statistically significant position effect for Vc (fig 2), the result of the large variance in this measurement, the clear trend of the supine position causing larger falls in Vc from baseline (−15.06% v −9.74%) compared with the seated position suggests that a gravity induced relocation of blood to the peripheral circulation is not the cause of the reduced central blood volume and therefore the depressed Vc.
Other possible hypotheses include compensatory shunting consequent on heavy exercise, as transient changes in venous tone in both pulmonary and systemic circulations direct blood flow away from the thorax to clear metabolic waste products from exercised muscles.10 This seems unlikely as the development of a depressed Vc after only 10 minutes of cycling at 25% Vo2max5 and the increasing magnitude of the impairment up to six hours after exercise6 do not reflect the time course of metabolic waste development and clearance respectively.
Increased thoracic electrical impedance, decreased levels of atrial natriuretic peptide,9 and decreased Vc all indicate a reduced central blood volume after exercise. The mechanism responsible for this cannot be determined from these data, although previous suggestions of passive redistribution of blood as the result of gravity can be discounted, suggesting that active vasoconstriction of the pulmonary vasculature and/or peripheral vasodilatation may be occurring after exercise.
ALVEOLAR CAPILLARY MEMBRANE DIFFUSING CAPACITY
After exercise, a decrease in Dm was observed (fig 3). This probably reflects an increase in the thickness of the alveolar capillary membrane, decreasing the rate of gas transfer between the alveolus and the red blood cells and/or a decrease in the available surface area for diffusion.
The decrease after exercise has been suggested to be an indicator of the development of acute pulmonary oedema. Direct attempts to quantify lung density and extravascular water content after exercise, using wet weight/dry weight ratio, indicator dilution, and computed tomography scanning techniques, have produced inconclusive results.3,25–27 The decreased Dm one hour after 10 minutes of low intensity exercise5 and the lack of effect on Pao2 and performance during repeated bouts of exercise11,28 suggest that physiologically significant injury to the alveolar capillary membrane is not the mechanism responsible for the decreased Dlco and Dm after short duration maximal exercise.
It is also possible that the less depressed Dm in the supine position is the result of increased pulmonary lymph flow, which could serve to remove any accumulated interstitial fluid. Cardiac output has been found to be linearly related to pulmonary lymph flow.17 Coates et al17 suggested that the increase in perfused microvascular surface area with exercise was the primary determinant for the increase in pulmonary lymph flow. If such a relation exists after exercise, then the increase in perfused microvascular surface area in the supine position29 may be expected to result in an increased rate of removal of any accumulated interstitial fluid.
The elevated Dm in the supine position compared with the upright seated position was probably due to gravity induced redistribution of blood from the distended capillaries in the lower zones of the lung, in the upright seated position, to the upper zones. This redistribution of blood could increase capillary recruitment and available surface area for diffusion in the supine position. The supine position decreased the ventilation to perfusion mismatch associated with an upright seated position.29 This increased uniformity in ventilation and perfusion in the supine position could increase Dm, through an increase in surface area available for gas transfer.
SUMMARY
This study confirms that Dlco is reduced after incremental cycling to volitional fatigue. It also indicates that the diffusion impairment is independent of measurement position. The change in Dlco appears to be primarily due to a decrease in Vc. The limited decrease in Dm in the supine position was probably due to redistribution of blood within the lung, as the result of gravity, increasing the surface area available for diffusion. The decrease in Vc has been previously attributed to reduced central blood volume. Although the mechanism for this reduction cannot be determined from these data, previous suggestions of passive relocation of blood to the periphery caused by gravity can be discounted, suggesting that active vasoconstriction of the pulmonary vasculature and/or peripheral vasodilatation may be occurring after exercise.
Take home message
The reduction in pulmonary diffusing capacity after exercise was not alleviated by altering body position, and was primarily due to a decrease in pulmonary capillary blood volume. The data indicate that the reduction in the latter was not due to passive relocation of blood, suggesting that active vasoconstriction of the pulmonary vasculature and/or peripheral vasodilatation may be occurring after exercise.