Article Text

Abstract
Objectives—To determine whether consecutive bouts of intense endurance exercise over a three week period alters serum concentrations of insulin-like growth factor I (IGF-I) and/or its binding proteins.
Methods—Seventeen professional cyclists (mean (SEM) Vo2max, 74.7 (2.1) ml/kg/min; age, 27 (1) years) competing in a three week tour race were selected as subjects. Blood samples were collected at each of the following time points: t0 (control, before the start of competition), t1 (end of first week), and t3 (end of third week). Serum levels of both total and free IGF-I and IGF binding proteins 1 and 3 (IGFBP-1 and IGFBP-3) were measured in each of the samples. Cortisol levels were measured in nine subjects.
Results—A significant (p<0.01) increase was found in total IGF-I and IGFBP-1 at both t1 and t3 compared with to (IGF-I: 110.9 (17.7), 186.8 (12.0), 196.9 (14.7) ng/ml at t0, t1, and t3 respectively; IGFBP-1: 54.6 (6.6), 80.6 (8.0), and 89.2 (7.9) ng/ml at t0, t1, and t3 respectively). A significant (p<0.01) decrease was noted in free IGF-I at t3 compared with both to and t1 (t0: 0.9 (0.1) ng/ml; t1: 0.9 (0.1) ng/ml; t3: 0.7 (0.1) ng/ml); in contrast, IGFBP-3 levels remained stable throughout the race.
Conclusions—It would appear that the increase in circulating levels of both IGF-I and its binding protein IGFBP-1 is a short term (one week) endocrine adaptation to endurance exercise. After three weeks of training, total IGF-I and IGFBP-1 remained stable, whereas free IGF-I fell below starting levels.
- cycling
- insulin-like growth factor
- exercise
- endurance
- binding proteins
Statistics from Altmetric.com
Take home message
Increased basal IGF-I is a short term (one week) adaptation to continuous bouts of highly demanding endurance exercise. However, this hormone shows no further increase after three weeks of exercise.
Insulin-like growth factor I (IGF-I) is a basic peptide composed of 70 amino acids and has a molecular mass of 7.5 kDa. With an amino acid sequence that is very similar to that of proinsulin, IGF-I plays an important role in the control of growth and metabolism. IGF-I expression is regulated by growth hormone (GH), and IGF-I acts as a feedback inhibitor at the pituitary level of GH release and/or GH gene transcription.1 Only 1% of IGF-I circulates freely in plasma; the remaining 99% is bound to carrier, or binding, proteins (BPs) of various types, the most important in humans being IGFBP-3 (>80%) and IGFBP-1. IGFBPs prolong the half life of IGF-I and act as autocrine and/or paracrine regulators of its biological activities by modulating the availability of IGF-I to cell surface receptors.2
Plasma IGF-I levels are regulated by factors that include the nutritional status of the subject, circulating levels of insulin and GH, and physical exercise.3 It has been established that short term exercise leads to a transitory increase in circulating IGF-I levels,4 but the IGF response to prolonged exercise and, in particular, its possible adaptations to training remain unknown. Several studies3, 5, 6 have shown that endurance exercise can provoke an increase in basal levels of total IGF-I. However, not all studies have been able to establish a significant increase in resting plasma levels as a result of periods of intense training.7 Evaluation of the possible changes in concentrations of IGF-I and its binding proteins may be of interest because it is thought that they may affect performance. Thus Koziris et al3 suggested possible effects of IGF-I in (a) the hypertrophy and maintenance of muscle mass in response to functional overload, (b) glucoregulation after training, and (c) the improvement in heart muscle contractility possibly resulting in increased systolic volume and cardiac output.
A further consideration is that changes in circulating levels of IGFBPs during exercise may modify the bioavailability of IGFs. IGFBP-1 is thought to be the major short term modulator of IGF bioavailability8 and is controlled by several factors such as insulin, glucocorticoids, and GH.9 The effects of prolonged training on serum IGFBP-1 have not been clearly established, although the results of several investigations have indicated increased concentrations after physical exercise.4, 10–12 IGFBP-3 has the ability to interact with cell surface receptors and increase the half life of IGF-I, potentiating its biological effects,13 but attempts to assess possible changes in serum IGFBP-3 concentration in relation to physical exercise have yielded controversial results.11, 14
Three week professional cycling competitions such as the Vuelta a España involve about 90 hours of intense exercise, during which cyclists cover about 3500 km with only one or two days of rest. Thus this type of competition constitutes an ideal model for the investigation of hormonal responses and adaptations to highly physiologically demanding exercise. Given the lack of studies performed on this type of athlete in terms of possible modifications in IGF-I levels during competition, the aim of this study was to determine whether three weeks of intense exercise (Vuelta a España 1999) could induce changes in resting serum levels of total IGF-I, free IGF-I, IGFBP-1, and IGFBP-3.
Methods
SUBJECTS
Seventeen cyclists from professional teams recognised by the International Cycling Union formed the study population. At the start of competition, the mean (SEM) age, height, and body mass of the participants were 27 (1) years, 179.6 (1.8) cm, and 66.9 (2.5) kg respectively.
About a month before participating in the study, all the subjects performed a ramp test on a cycle ergometer for determination of maximal oxygen uptake (Vo2max). After a five minute warm up at 50 W, the workload was increased by 25 W/min until exhaustion. All the subjects had previous experience of this type of protocol, which has often been used for the physiological evaluation of professional cyclists. Vo2max averaged 74.7 ml/kg/min.
Mean percentage body fat was estimated at 8.1 (0.1)% using a skinfold method described elsewhere.15 All the subjects were in good health, as established by a normal physical examination (including electrocardiographic and echocardiographic evaluation) within the previous months. Mean daily food intake during this type of competition is about 840 g carbohydrate, 200 g protein, and 158 g fat corresponding to about 23.5 MJ.16 No participant had a personal or family history of endocrine or metabolic disorders. No exogenous substances were detected during routine “anti-blood doping” analysis. Informed consent was obtained from each participant in accordance with the regulations of the Universidad Complutense, Madrid.
CONDITIONS BEFORE THE RACE
All the subjects followed the same lifestyle during the week before the start of the race. Their diet was standardised (breakfast between 0930 and 1000 after nine hours of sleep; lunch at about 1400; dinner at 2030), and carbohydrate intake averaged 500–600 g/day. All the subjects had covered an average of about 30 000 km from the start of the training season (December). They underwent a tapering week before the start of the race (3–4 h/day of easy to moderate cycling, most of which was performed at heart rates below 160 beats/min).
EXERCISE CHARACTERISTICS (VUELTA A ESPAÑA 1999)
A distance of 3518 km was covered in 21 stages over the three week period which included one day of rest. This distance was covered by the overall winner in about 90 hours (at an average speed of about 39 km/h). The mean distance covered per daily stage was 168 (11) km at altitudes of about 0–2200 m above sea level.
PROTOCOL
Blood samples were drawn from each participant just before the start of competition (control t0) and at the end of the first (t1) and third (t3) week of competition. All blood sampling was performed between 0900 and 0930 after an overnight fast and nine hours of sleep. Each subject was lying supine on a bed during blood collection. The blood was centrifuged and serum was stored immediately at −30°C until tested.
Immediately before blood collection, each cyclist was weighed and percentage fat was determined.
ANALYTICAL METHODS
Total IGF-I in serum
Serum concentrations of total IGF-I were estimated by double antibody radioimmunoassay, after removal of serum IGFBPs by acid-ethanol extraction.17 To confirm the removal of IGFBPs, extracted and non-extracted serum fractions were incubated with 125I-IGF-I at 4°C for 24 hours. Dextran charcoal was used to separate the bound and free tracers. Extracted fractions contained 98% free IGF-I whereas non-extracted fractions contained only 42% free IGF-I. The IGF-I antiserum (UB2-495) was a gift from Drs Underwood and Van Wisk distributed by the Hormone Distribution Program of the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK) through the National Hormone and Pituitary Program. Levels of IGF-I were expressed in relation to the IGF-I standard A52-EPD-186 (Eli Lilly). Test sensitivity was 28 ng/ml and the intra-assay coefficient of variation was 8%. All samples were run in the same assay.
Free IGF-I, IGFBP-1, and IGFBP-3 in serum
Serum concentrations of free IGF-I, IGFBP-3, and IGFBP-1 were determined in duplicate by immunoradiometric assay using commercially available automated kits (Diagnostic System Laboratories, Webster, Texas, USA). Assay sensitivities were 30 pg/ml for free IGF-I, 330 pg/ml for IGFBP-1, and 500 pg/ml for IGFBP-3. Intra-assay coefficients of variation were 5%, 4.6%, and 3.2% respectively.
Serum cortisol
Cortisol was determined in nine subjects using a commercial automated enzyme immunoassay kit (enzyme linked fluorescent assay; VIDAS, Biomérieux, France). Reference values were 60.0–285.0 ng/ml. The assay sensitivity and intra-assay coefficient of variation were 3.0 ng/ml and <16% respectively.
Data were corrected for percentage variations in plasma volume according to a method described elsewhere.18
STATISTICAL ANALYSIS
Results are expressed as means (SEM). Once the Kolmogorov-Smirnov test was applied to establish a gaussian (normal) distribution of data, basal (morning) levels of total IGF-I, free IGF-I, IGFBP-1, IGFBP-3, and cortisol were compared by repeated measures analysis of variance. The Newman-Keul's test was used post hoc to identify significant differences. Finally, correlations were calculated between free IGF-I and IGFBP-1 levels (Pearson product moment correlation). p<0.05 was taken to denote statistical significance.
Results
Mean body mass fell at the end of competition with respect to starting values (65.8 (2.2) v 66.9 (2.5) kg; p<0.05); no change was recorded after the first week (66.2 (2.5) kg). Percentage body fat values did not vary significantly (p>0.05) throughout the competition (8.2 (0.1)%, 8.2 (0.1)%, and 8.1 (0.1)% for t0, t1, and t3 respectively).
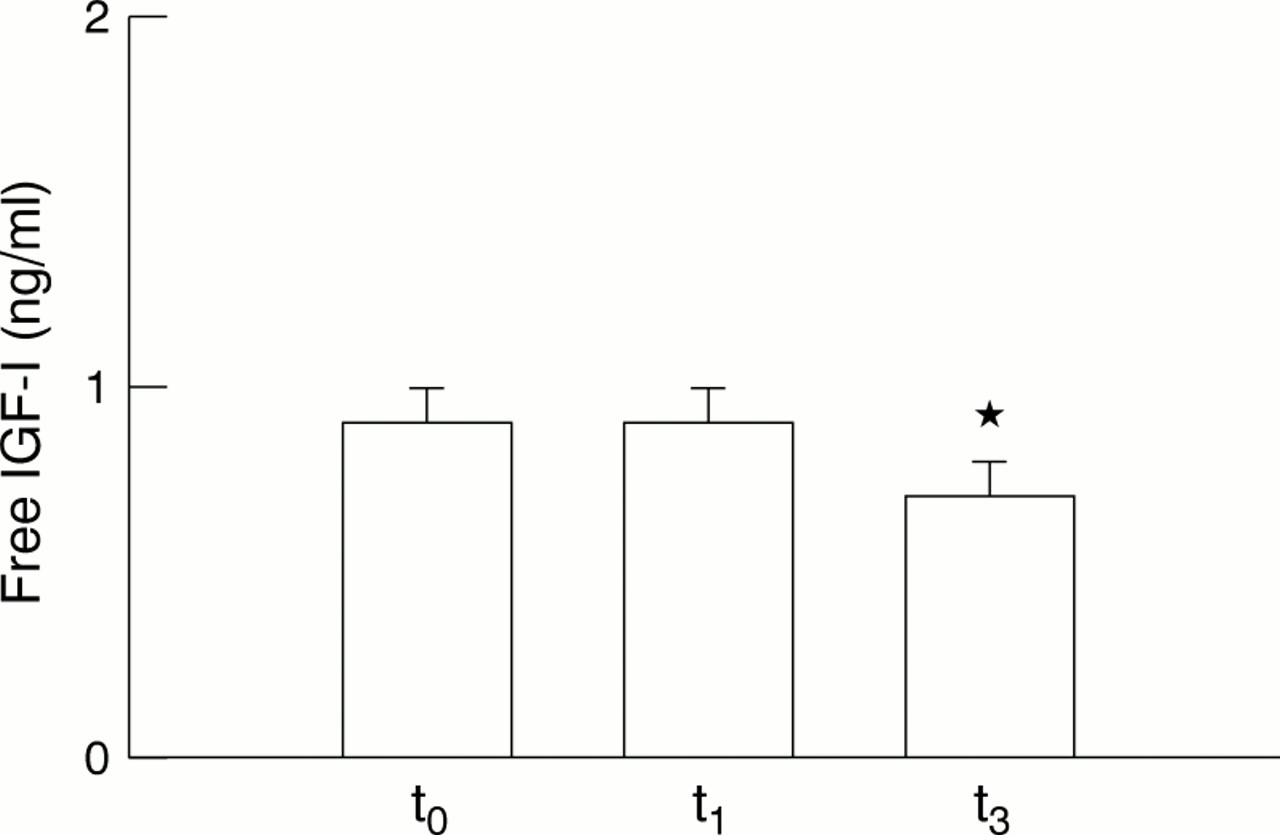
Mean values of total IGF-I recorded at t0 were significantly lower than those recorded at t1 and t3 (p<0.01) (fig 1). These values correspond to increases of 68.4% (t1) and 77.5 % (t3) above starting levels (t0). Total IGF-I levels recorded at t1 and t3 were not significantly different (p>0.05). There were no changes in free IGF-I levels between t0 and t1 (p>0.05) (fig 2). However, a significant fall (p<0.01) was shown at t3 with respect to levels noted at t0 (−31.9%) and t1 (−25.0%).

Comparison of resting serum concentrations of total insulin-like growth factor I (IGF-I). Values are expressed as mean (SEM). *p<0.01 for the comparison t0 v both t1 and t3.
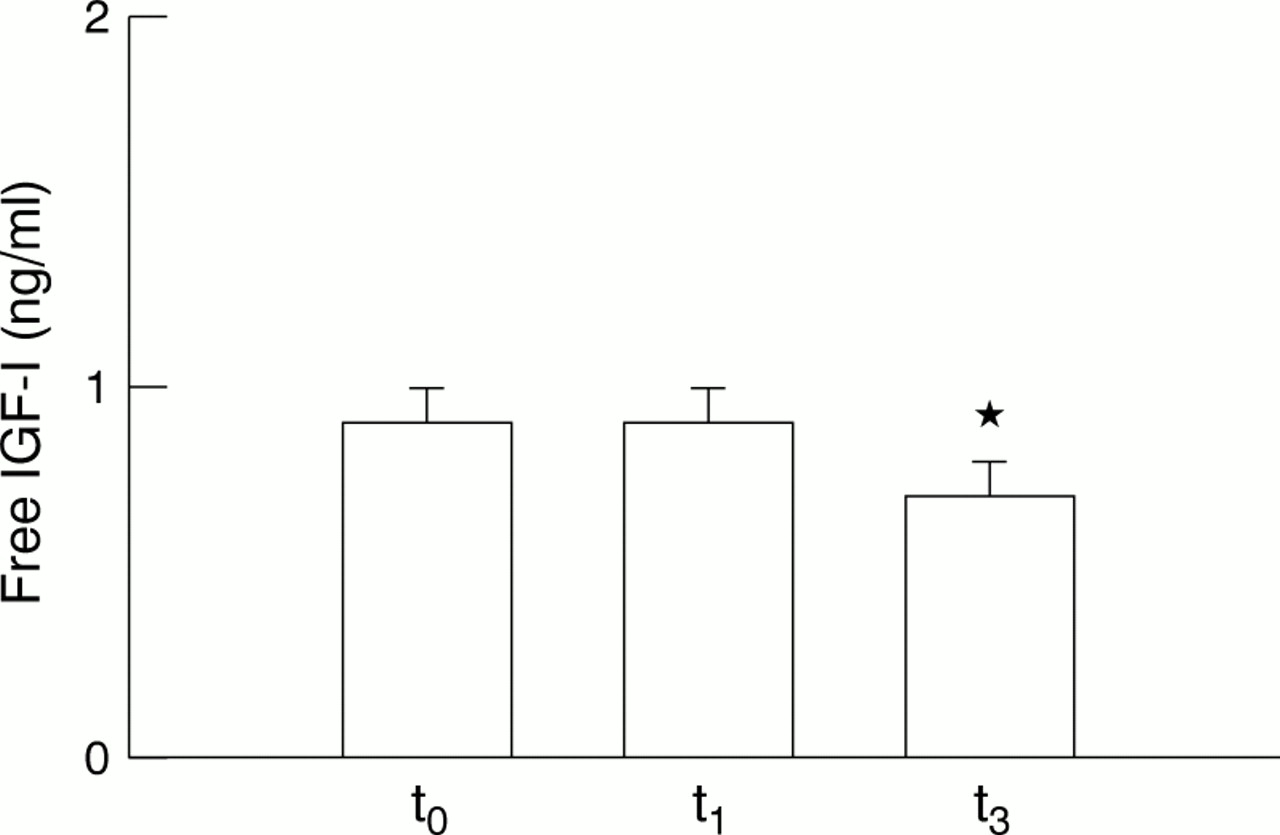
Comparison of resting serum concentrations of free insulin-like growth factor I (IGF-I). Values are expressed as mean (SEM). *p<0.01 for the comparison t3 v both t0 and t1.
Average values of IGFBP-1 were significantly higher at both t1 and t3 compared with t0 (p<0.01) (fig 3), representing increases of 47.6% (t1) and 63.3% (t3) over starting levels (t0). No significant differences were found between t1 and t3 (p>0.05). On the other hand, no significant differences (p>0.05) were observed in serum IGFBP-3 levels throughout the study period (t0, t1, and t3) (fig 4).

Comparison of resting serum concentrations of insulin-like growth factor binding protein 1 (IGFBP-1). Values are expressed as mean (SEM). *p<0.01 for the comparison t0 v both t1 and t3.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Serum concentrations of insulin-like growth factor binding protein 3 (IGFBP-3) during competition. Values are expressed as mean (SEM). There were no significant differences between means.
Levels of serum cortisol at t0 and t1 did not vary significantly (217.8 (19.2) and 170.3 (12.9) mIU/ml respectively; p>0.05), whereas significantly (p<0.01) lower levels were recorded at t3 (137.2 (24.6) mIU/ml) with respect to t0 (−37.0%) and t1 (−19.4%).
Finally, levels of free IGF-I correlated significantly with the corresponding IGFBP-1 levels (r = −0.47; p<0.01).
Discussion
The main finding of this study was the appreciable increase in total IGF-I and IGFBP-1 observed during the first part of this prolonged endurance competition, and the significant fall in free IGF-I recorded at the end of the three week race. The physiological significance of adaptations in IGF-I levels induced by endurance cycling is yet to be established, although IGF-I may be involved in the processes of muscle mass maintenance associated with severe functional overload19, 20 and enhanced myocardial contractility.20, 21
The increase in serum total IGF-I and IGFBP-1 observed after the first week of competition is in agreement with previous findings3, 6, 22 that also indicate this type of kinetics after a brief period of intense exercise. Roelen et al6 observed increased plasma levels of IGF-I and IGFBP-1 after two weeks of cycle training. Koziris et al3 reported positive effects of training on resting serum levels of total IFG-I in swimmers. The great intensity and duration of exercise performed by subjects in this study may explain the early rise (first week) in serum total IGF-I. The studies mentioned above did not explore possible changes during the first week of exercise. Given that neither subject body mass nor percentage body fat showed significant change at t1, it is unlikely that the increase in IGF-I recorded over the first week are due to a change in the energy status of our cyclists. It is known that GH induces short term increases in tissue IGF-I production, leading to increased serum levels,23 and that physical exertion stimulates GH release in the circulation.24 Thus it may be that increased GH secretion during the competition could have effects on serum IGF-I levels. However, some authors10, 25, 26 have noted a long term dissociation between increased GH and enhanced IGF-I levels. The design of our study did not permit us to speculate on the kinetics of GH during this type of competition.
The increase in serum IGFBP-1 concentration may have an important role in glucoregulation, controlling the bioavailability and bioactivity of IGF, especially as the half life of the IGF-I/IGFBP-1 complex has been shown to be much shorter than that of IGF-I/IGFBP-3.27 During the first week of competition, our cyclists showed significantly increased serum IGFBP-1 levels, consistent with the findings of previous studies.4, 10–12 Hopkins et al10 detected significantly increased IGFBP-1 levels after prolonged exercise despite maintenance of stable glucose and insulin concentrations, suggesting that other factors are involved in the increased production of IGFBP-1. It has been proposed that the rise in serum IGFBP-1 that occurs in response to exercise in some way stimulates sequestration of free IGF-I and thereby prevents insulin-like action.28 This could explain why free IGF-I was unmodified during the first week of competition in our cyclists. Further, IGFBP-1 has been shown to act as a potent inhibitor of in vivo IGF-I bioactivity,29, 30 possibly through the regulation of free IGF-I levels,31 given the inverse relation between serum IGFBP-1 and free IGF-I.32 We also noted this negative relation, which would appear to support the inhibitory activity of IGFBP-1 on free IGF-I. Investigations performed to evaluate possible changes in IGFBP-3 after physical exercise have provided equivocal results.11, 14 In this study, we detected no change in IGFBP-3 throughout the competition, and, despite the fact that we did not examine the system's proteolytic capacity of IGFBP-3, it would seem that this activity was not stimulated during the initial days of competition in the cyclists analysed.
The behaviour of IGF-I and IGFBP-1 showed considerable variation at three weeks of competition. Both total IGF-I and IGFBP-1 failed to show a continued increase but remained stable with respect to values recorded after one week. However, values in the third week were higher than those before the competition. It has been observed that high intensity training or prolonged endurance activities provoke a long term decrease in IGF-I levels.33–35 Our subjects performed exercise of greater intensity during the last two weeks of competition, because the final stages of competition included mountain passes resulting in higher metabolic demand.36 This was reflected in the fall in body weight recorded at the end of the competition. It has been established that reduced energy intake leads to reduced IGF-I concentrations. Furthermore, it is possible that increased protein degradation for energetic ends (explaining the fall in body mass and maintenance of percentage fat) may have provoked the arrest observed in the increase in IGF-I concentration in the cyclists examined.37, 38 On the other hand, as IGFBP-1 production is regulated by glucocorticoids,9 its stabilisation noted in the third week with respect to the first may be related to the fall in blood cortisol detected in these athletes during the final days of competition. This decrease in cortisol may be caused by a possible depletion of the adrenal gland as a result of the chronic increase in secretion demand. It is also possible that, owing to the greater tissue requirement for IGF-I in the active muscle subjected to intense exercise over several days, the IGFBP-1 complex crosses the endothelium39 directing IGF-I from the general circulation towards muscle receptors. Hopkins et al10 proposed that, during prolonged exercise, IGFBP-1 directs circulating IGF to skeletal muscle, in which IGF-I receptors are abundant and IGF facilitates the consumption of glucose. Further, increased IGF-I secreted by the liver would be rapidly sequestered by tissues in an attempt to maintain homoeostasis in the systemic circulation; this would prevent IGF-I inhibiting GH secretion by acting on the pituitary and hypothalamus. This sequence of events could explain the reduction in free IGF-I observed here in the blood sample taken at three weeks as well as the stable serum concentrations of IGFBP-1 and total IGF-I. Furthermore, Suikkari et al12 found that, when the energetic cost exceeds energy consumption, a fall in free IGF-I levels occurs.
Before drawing definitive conclusions, it must be borne in mind that there is support for increased IGF-I production in muscle tissue. Indeed, Eliakim et al40 reported an increase in the total production of IGF-I in response to exercise in rats after five days of training, despite a lack of increased serum IGF-I levels. This lack of change in serum levels suggests that serum IGF-I level may not be an adequate marker of GH-IGF-I system activity. In an attempt to explain the observation of catabolic-type responses to five weeks of exercise training, Eliakim et al33 suggested that local muscle IGF-I may actually increase as circulating IGF-I falls, reflecting an important mechanistic role for autocrine/paracrine effects of IGF-I in response to training. Considering the postulated physiological advantages that the rise in serum IGF-I concentration may have in endurance exercise performance, our findings should prompt future studies of the adaptation processes of this peptide and its carrier proteins.
In conclusion, the results of this investigation suggest a short term increase in resting serum IGF-I and IGFBP-1 levels (one week) in top level cyclists during prolonged endurance competition, with long term (three weeks) stabilisation of the levels accompanied by a significant reduction in free IGF-I and no change in IGFBP-3. The metabolic consequences of these alterations on physical performance need further investigation. Indeed, we can hypothesise an increased demand for IGF-I by tissues that are active during this type of exercise.
Acknowledgments
This study was financed by the Asociación Deportiva Banesto. Thanks are due to Ana Burton for translating the manuscript.
Take home message
Increased basal IGF-I is a short term (one week) adaptation to continuous bouts of highly demanding endurance exercise. However, this hormone shows no further increase after three weeks of exercise.