Article Text

Abstract
Objectives—To evaluate the hormonal response to strenuous endurance exercise performed by elite athletes.
Methods—Nine professional cyclists (mean (SD) age 28 (1) years; mean (SD) Vo2max 75.3 (2.3) ml/kg/min) who participated in a three week tour race (Vuelta a España 1999) were selected as subjects. Morning urinary levels of 6-sulphatoxymelatonin (aMT6s) and morning serum levels of testosterone, follicle stimulating (FSH), luteinising hormone (LH), and cortisol were measured in each subject at t0 (before the competition), t1 (end of first week), t2 (end of second week), and t3 (end of third week). Urine samples of aMT6s were also evaluated in the evening at t0, t1, t2, and t3.
Results—Mean urinary aMT6s levels had increased significantly (p<0.01) during the day after each stage (1091 (33) v 683 (68) ng/ml at t1; 955 (19) v 473 (53) ng/ml at t2; 647 (61) v 337 (47) ng/ml at t3). Both morning and evening aMT6s levels decreased significantly during the study. A similar pattern was observed for morning serum levels of cortisol and testosterone.
Conclusions—The results suggest that the basal activity of the pineal gland, adrenal glands, and testis may be decreased after consecutive days of intense, long term exercise.
- melatonin
- gonadotrophins
- testosterone
- cortisol
- endurance exercise
Take home message
The night time activity of the pineal gland seems to be attenuated after consecutive days of vigorous daytime exercise, namely three week tour races for professional cyclists. A similar state of exhaustion seems to occur in the adrenal glands and testis.
Statistics from Altmetric.com
Melatonin is secreted at night by the pineal gland in a rhythmical fashion.1 Its secretion and hence rhythm is directly decreased by bright light.1 When administered exogenously, melatonin can alter the timing of the internal body clock.2 Some studies have shown that strenuous exercise—for example, long distance running—during the day can increase melatonin levels, particularly in women.3–,8 The levels return to basal values after 30–60 minutes of recovery from exertion.3,6 These findings, however, have not been corroborated by other authors using either women or men as subjects.9,10 Further, the reported effects of exercise on melatonin secretion may have been influenced by natural bright light. On the other hand, the acute increase in circulating melatonin that can occur after an exercise bout has been found to be attenuated by regular, vigorous training.5 Whether the rise in plasma melatonin induced by exercise has physiological relevance is not known.
It is well documented that exhaustive exercise can lead to activation of several “stress hormones”. For instance, prolonged exercise can maximally stimulate the hypothalamic-pituitary-adrenal (HPA) axis. During intense prolonged exercise, adrenocorticotrophin (ACTH) concentrations show a significant increase, which in turn results in appreciable release of cortisol.11 Such responses are exaggerated under adverse conditions such as increases in body temperature or psychological stress.12 Basal cortisol levels may also be increased after long term training13 or overtraining.14
Extensive research has been conducted on the responses and adaptations of the hypothalamic-pituitary-testicular (HPT) axis to endurance exercise. During prolonged exercise, the response of testosterone appears quite variable: both increases and decreases have been reported.11,12 Basal levels of testosterone, on the other hand, can show subclinical decreases in endurance trained humans.15–,20 Moreover, the higher the training loads, the lower the basal testosterone in those showing training induced decreases in this hormone.11 More research is, however, needed on the influence of exercise and training on the gonadotrophins, luteinising hormone (LH) and follicle stimulating hormone (FSH). The response of both hormones to prolonged exercise does show considerable variability, with significant increases, decreases, or no changes at all being reported.12,20,21 The resting pulsatile characteristics of LH seem to be more consistently affected after training,22,23 than its overall basal levels, which can decrease,19 or remain unchanged.16,18,24 Although the behaviour of FSH is even more inconsistent, no major alteration is usually found in this hormone with training.16,18,24
Professional cycling races such as the Vuelta a España last 22 days (about 90 hours of competition), during which participants must cover about 3500 km with only one or two days of rest. Despite the long duration of the daily stages (four to five hours and 150–200 km), exercise intensity is often high—that is, anaerobic metabolism is likely to prevail during the more physically taxing events (mountain passes, time trials, sprints, “breakaways”, etc).25,26 To the best of our knowledge, no other investigation has been designed to explore the endocrine response provoked by such an extreme endurance competition (combining both long exercise duration and high intensity bouts). In addition, no previous report has evaluated the melatonin response to exercise in subjects of such high fitness level. Thus, the aim of this investigation was to evaluate the endocrine response of professional cyclists during a three week tour competition, the 1999 Vuelta a España. The levels of several circulating hormones known to play a role in the body responses to exercise—that is, hormones of the HPT and HPA axes11—and their possible relation to melatonin were determined.
Methods
SUBJECTS
Nine male cyclists from one of the world's leading professional teams were recruited to participate in the investigation. None of the subjects had travelled between time zones before the start of the race. All are highly competitive in the professional category, as shown by their outstanding competition results: two were in the top 15 (including 1st and 2nd) to complete the Giro d'Italia, Vuelta a España, and Tour de France 1995–1999.
Their age, height, body mass, and maximal oxygen uptake (Vo2max) were 28 (1) years, 180.8 (1.9) cm, 67.1 (2.8) kg, and 75.3 (2.3) ml/kg/min respectively (mean (SEM) at the start of the Vuelta a España. Percentage body fat was estimated at 8.2 (0.2)% using a skinfold method described elsewhere.27 All the subjects were in good health as established by a normal physical examination (including electrocardiographic and echocardiographic evaluation) within the previous months. They had normal endocrine and metabolic functions and no family history of diabetes mellitus. Written informed consent was obtained from each subject in accordance with the regulations of the Universidad Complutense, Madrid.
VUELTA A ESPAÑA 1999: MAIN CHARACTERISTICS
The race started in late August at the Spanish Mediterranean coast and finished in mid September. It included 21 consecutive daily stages with only one day of rest. During this three week period, 3518 km were covered over a large area of Spain (excluding the southern part). The total time spent racing by the overall winner was about 90 hours (average speed about 39 km/h). The mean distance covered per daily stage was 168 (11) km at altitudes of about 0–2200 m above sea level. Most daily stages started at about 1200 and finished between 1630 and 1730. Temperatures ranged from 18 to 30°C during the stages. Most days were sunny, including those on which we collected blood and urine samples (estimated light conditions 5000–6000 lux).
To maintain circadian rhythms, nutrition, hydration, timing of food intake, and sleep were kept constant throughout the three week period, which is normal practice in cycling competitions.28 Mean daily food intake during this type of competition is about 840 g carbohydrate, 200 g protein, and 158 g fat corresponding to about 23.5 MJ.28
EXERCISE TEST
Each subject performed an incremental exercise test a few weeks before participating in the race so that reference heart rates corresponding to the lactate threshold (LT) could be determined. The tests were performed on a bicycle ergometer following a ramp protocol until exhaustion (workload increases of 25 W/min), as described elsewhere.25 Heart rate (beats/min) was continuously recorded during the tests with a telemeter (Polar Xtrainer Plus, Polar Electro, Oy, Finland). Capillary blood samples were taken from fingertips (25 μl) every two minutes and immediately after exercise with an electroenzymatic analyser (YSI 1500 Sport; Yellow Springs Instruments, Yellow Springs, Ohio, USA). The exercise intensity corresponding to the LT was determined as described by Weltman et al29 and averaged about 75% of the subjects' Vo2max.
ANALYSIS OF EXERCISE INTENSITY DURING THE RACE
All subjects were familiar with the use of the telemeter to monitor heart rate during the whole of each stage of the Vuelta a España. Recorded data were analysed using a computer program (Polar Heart Rate Analysis Software 5.03; Polar Electro) that allows the user to select reference heart rate values and to establish different levels of exercise intensity. We quantified the total time spent during each daily stage at an exercise intensity above LT. Exercise performed above LT can be defined as “intense” exercise—that is, with increasing involvement of anaerobic metabolism in total energy production.26
HORMONE MEASUREMENTS
Four blood samples were withdrawn from each participant as follows: at t0 (before the competition; control), t1 (end of first week), t2 (end of second week), and t3 (end of third week). All blood sampling was performed (a) after an overnight fast and nine hours of sleep (from 2400 to 0900) in the dark (light intensity <0.2 lux), and (b) between 0900 and 0930 under dim light conditions (light intensity about 100 lux).
Subjects were made comfortable for at least 10 minutes before blood collection. A blood sample was then withdrawn by clean venepuncture from each subject for the determination of resting levels of the following hormones: LH, FSH, testosterone, and cortisol. The samples were allowed to clot at room temperature and then centrifuged at 2000 g for 20 minutes at 4°C. Separated serum was divided into aliquots and stored at 2–8°C for the determination of LH, FSH, and cortisol, and at −20°C for testosterone determination. Serum levels of LH, FSH, and cortisol were quantified within 10 hours of sample collection using a commercial automated enzyme immunoassay kit (enzyme linked fluorescent assay; VIDAS; Biomérieux, Lyon, France). The interassay variability was <6.6%, <3.0%, and <11.5% for LH, FSH, and cortisol respectively. The intra-assay variability was <6.3%, <4.3%, and <16.2% for LH, FSH, and cortisol respectively. Assay sensitivity was 0.04 IU/l for LH, 0.03 IU/l for FSH, and 3.0 ng/ml for cortisol. Reference values were 1.1–7.0 IU/l for LH, 1.7–12.0 IU/l for FSH, and 6–285 ng/ml for cortisol. Levels of testosterone were estimated using a solid phase 125I radioimmunoassay kit (Diagnostic Products, Los Angeles, California, USA). The interassay and intra-assay coefficients of variation were both equal to 5.8%, whereas assay sensitivity was 0.1 ng/ml. Reference values for testosterone were 2.6–15.9 ng/ml.
Two urine samples each of volume 150 ml were collected before (0900–0930) and after (2000–2030) the corresponding daily stage from each participant at t1, t2, and t3. Two urine samples (150 ml) were also collected in the same manner at to, before competition started. Aliquots of volume 1 ml from each sample were frozen and stored at −20°C until measurement of urinary 6-sulphatoxymelatonin (aMT6s) in duplicate by radioimmunoassay (RIA) (Stockgrand Ltd, Surrey, UK). This method is a modification30 of the RIA reported by Arendt et al.31125I-aMT6s radiolabelled freeze dried sheep antiserum and charcoal stripped (aMT6s-free) urine were used to separate antibody bound aMT6s from the free fraction. The sensitivity of the assay was 0.5 ng/ml. The intra-assay coefficient of variation was 5.2%. Studies in humans have reported 10 times lower levels of aMT6s in urine than blood,32 and urinary excretion of aMT6s has been shown to be a valid measure of melatonin rhythm and melatonin secretion into the blood in humans32–,34 and experimental animals.35
Subjects were weighed in the morning of t0, t1, t2, and t3, before blood and urine sampling. To minimise the confounding effects of exercise induced dehydration on urine concentration in the evening of t1, t2, and t3, subjects were (a) instructed to drink at least 1.5 litres between the end of the stage and urine collection, and (b) weighed again before collection of urine samples during the evening. To minimise any possible effect of changes in plasma volume on morning levels of aMT6s, FSH, LH, testosterone, and cortisol over the three week period—that is, variations in hydration status or training induced haemodilution throughout the tour race—we calculated the percentage changes in plasma volume between morning blood samples collected at t1, t2, and t3 compared with to values using a method that requires determination of packed cell volume (%) and haemoglobin concentration (g/dl) in each blood sample.36
STATISTICAL ANALYSIS
Once the Kolmogorov-Smirnov test was applied to establish a Gaussian distribution of data, morning levels of FSH, LH, cortisol, testosterone, and aMT6s recorded throughout the three week period (from t0 to t3) were compared by repeated measures analysis of variance. The same statistical analysis was used to compare levels of aMT6s recorded in the evening throughout the study (from t0 to t3). The Newman-Keuls test was used post hoc to identify significant differences. We used Student's t test for paired data to compare mean levels of aMT6s recorded in the morning and those recorded in the evening at t0, t1, t2, and t3—that is, morning v evening at to, morning v evening at t1, etc). Finally, correlation coefficients between the different hormones were also calculated.
Results are expressed as mean (SEM) and p<0.05 was taken to denote significance.
Results
The mean total duration of the Vuelta a España for our subjects was 90 hours 51 minutes. The mean duration of each stage was 242 (23) minutes (about four hours), and the total time spent per stage at an exercise intensity above LT (>75% of Vo2max) averaged 71 (7) minutes.
The mean body mass of the subjects measured in the morning remained stable throughout the three week period—that is, only a 1.6% decrease from t0 to t3 (t0 67.1 (2.8) kg; t1 66.4 (2.5) kg; t2 66.1 (2.3) kg; t3 66.0 (2.2) kg). Plasma volume shifts throughout the study (t1, t2, t3) compared with before the tour (t0) were less than 2.0% in all the subjects. Finally, the percentage change in body mass between morning and evening was −0.1% at t1, + 0.6% at t2, and + 0.9% at t3, which indirectly confirms that subjects were euhydrated before urine sampling in the evening.
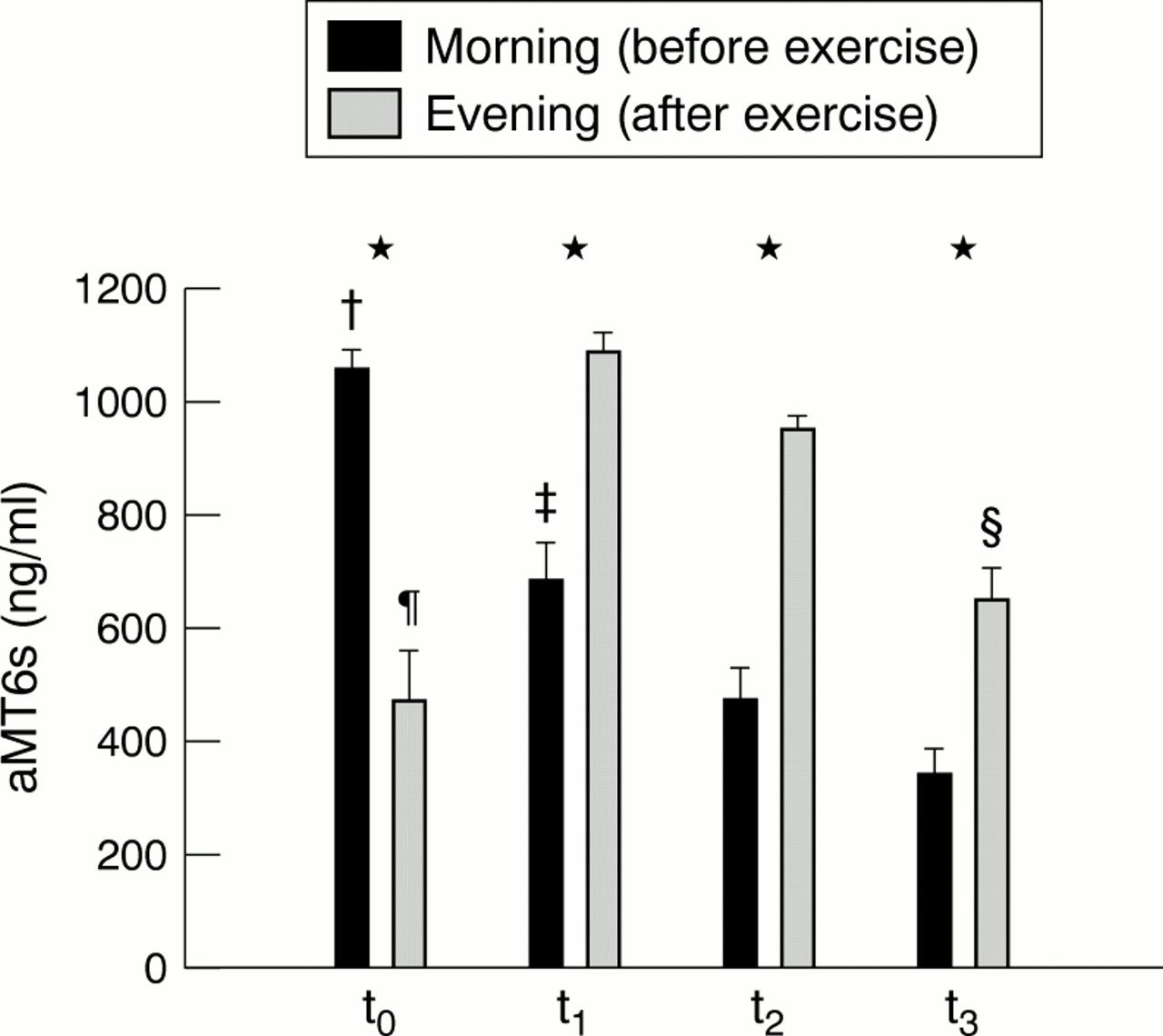
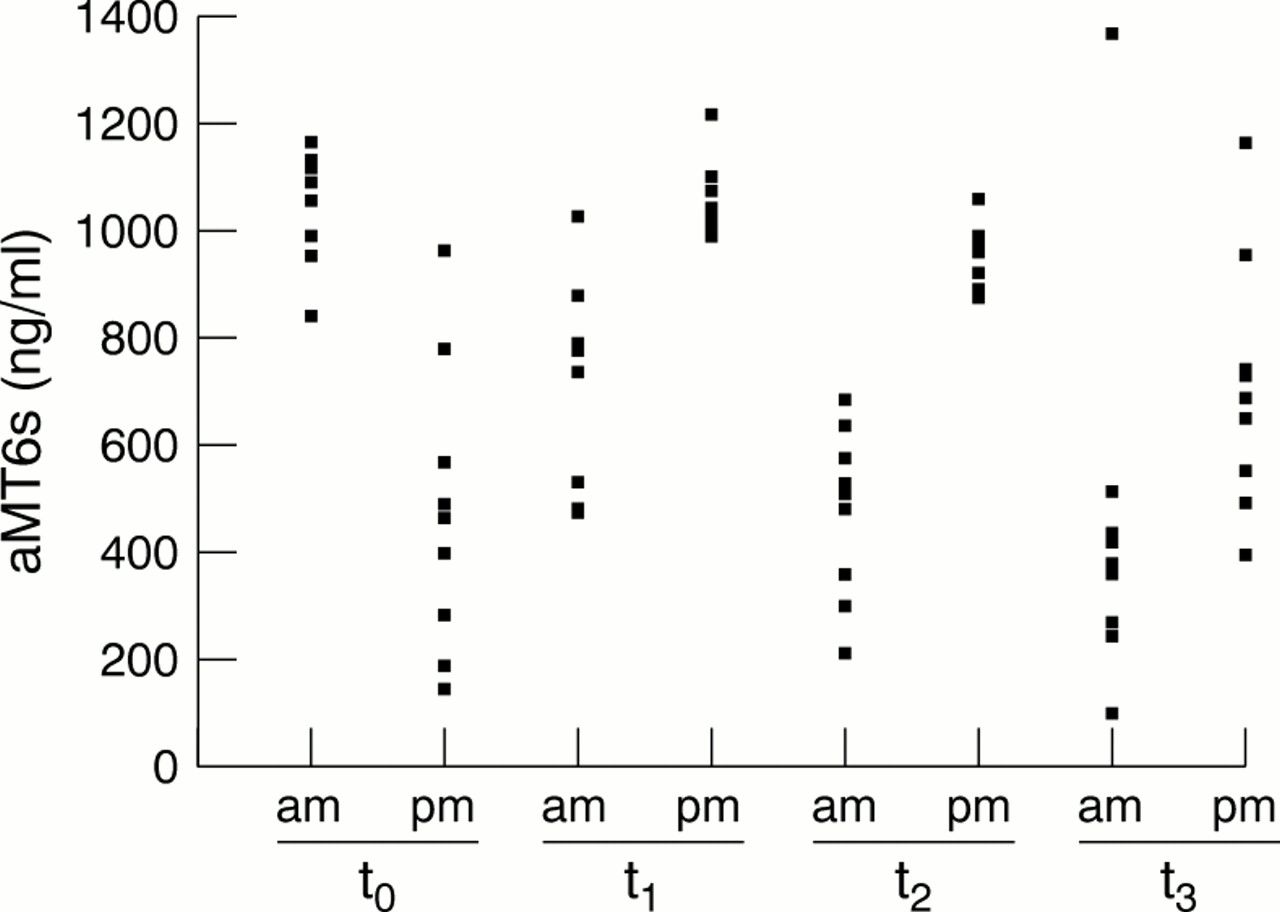
Figures 1 and 2⇓⇓ show mean and individual values respectively of urinary aMT6s obtained during the study. Before competition started (to), morning levels of aMT6s were significantly higher (p<0.01) than those collected in the evening. In contrast, at t1, t2, and t3, levels of aMT6s after exercise were increased (p<0.01). Levels of aMT6s measured in the morning and evening both decreased during the study.

Morning (0900—0930) and evening (2000—2030) levels of 6-sulphatoxymelatonin (aMT6s) in urine before the competition (t0) and at the end of the first (t1), second (t2), and third (t3) week. Values are expressed as mean (SEM). The following significant differences were found: *p<0.01 for morning v evening; †p<0.01 for t0 morning v t1, t2, and t3 morning; ‡p<0.01 for t1 morning v t2 and t3 morning; §p<0.01 for t3 evening v t1 and t2 evening, and p<0.05 for t3 evening v t0 evening; ¶p<0.01 for t0 evening v t1 and t2 evening (p<0.01).

Individual values of 6-sulphatoxymelatonin (aMT6s) concentration in urine before the competition (t0) and at the end of the first (t1), second (t2), and third (t3) week. am, Morning, before exercise; pm, evening, after exercise.
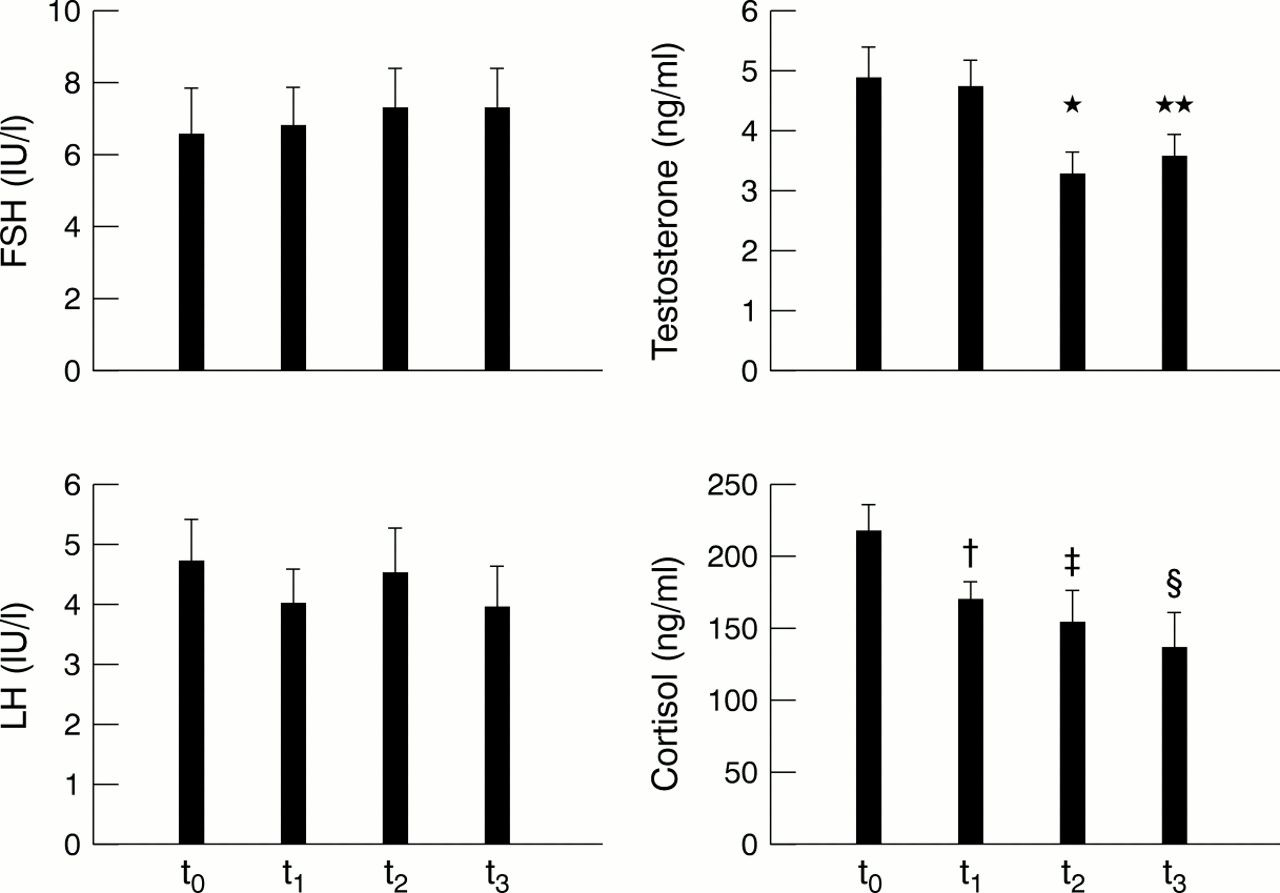
No significant differences were noted in LH or FSH levels throughout the race (fig 3⇓). Serum testosterone and cortisol, in contrast, decreased throughout the race.

{kind=link}
{kind=link}
{kind=link}
Morning serum levels of follicle stimulating hormone (FSH), luteinising hormone (LH), testosterone, and cortisol before the competition (t0) and at the end of the first (t1), second (t2), and third (t3) week. Data are mean (SEM). There were no significant differences between means for LH and FSH. The following significant differences were found for testosterone and cortisol: *p<0.01 for t2 v t0 and t1; **p<0.01 for t3 v t0 and t1; †p<0.05 for t1 v t0; ‡p<0.01 for t2 v t0; §p<0.01 for t3 v t0.
Finally, no significant correlations existed between the different hormones.
Discussion
To the best of our knowledge, this is the first attempt to evaluate the endocrine response to many consecutive days of highly demanding exercise in elite endurance athletes. No previous report has evaluated the melatonin response to exercise in subjects of such high fitness level. In addition, to date there are no studies showing a significant decrease in morning (before exercise) levels of hormones of both the HPT and HPA axes after only three weeks of strenuous exercise.
A field study such as this is not without limitations. For instance, exercise induced dehydration could artificially alter aMT6s levels in urine between morning and evening. However, the body mass of the subjects before evening sampling was very similar to that measured during the morning. Indeed, it is common practice for professional cyclists to drink considerable amounts during three week stage races—that is, 6.7 litres each per 24 hour period, and as much as 4 litres of this during the stage.37 Furthermore, our subjects had consumed at least 1.5 litres of fluids between the end of the stage (about 1730) and urine collection (2000–2030).
ACUTE INCREASES IN AMT6S WITH DAYTIME EXERCISE
Our findings showing significantly higher aMT6s secretion after the stages at the end of the first, second, and third week compared with before the competition are in agreement with previous studies which have shown that strenuous and less intense exercise protocols—for example, long distance running—carried out during the day can increase melatonin levels, particularly in women.3–,8 These findings, however, have not been corroborated by other authors.9,10 Taken together, the present data and those of previous research3–,8 suggest that moderate to intense, long lasting exercise—that is, several hours at more than 50% of Vo2max—during the day may stimulate melatonin secretion, at least partly.
Although the physiological mechanisms involved in this stimulatory effect have yet to be determined, several have been postulated. Studies in which both circulating catecholamines and melatonin were measured during the day in rats showed that peripheral catecholamines released by certain stressful stimuli may promote pineal melatonin production and increase serum melatonin levels.3–,6 In contrast, adrenalectomy blocked the increase in rat pineal melatonin content brought about by daytime physical immobilisation stress and insulin induced hypoglycaemia.3–,5 One could expect in our subjects a catecholamine induced rise in serum melatonin after most daily stages, given that (a) the contribution of high intensity exercise—that is, above LT or 75% of Vo2max—to the total exercise duration of each daily stage (about four hours) was not negligible (71 minutes), and (b) it has been well documented that plasma catecholamine concentrations increase considerably at intensities above the LT.38 The aforementioned mechanism, on the other hand, is probably not operative until a certain duration of exercise is reached (several hours), given that there is no significant storage of melatonin in the pineal gland.39 In any case, some limiting factors, namely time of day and hence environmental light, should be taken into consideration when interpreting the results of studies on the effects of daytime exercise on melatonin secretion. Our finding of an increase in aMT6s after exercise must be interpreted with caution because it does not necessarily imply a direct stimulating effect of daytime exercise on melatonin secretion. Indeed, the profile of urinary aMT6s secretion at t1, t2, and t3 is similar to that expected in healthy humans performing no exercise40—that is, a progressive increase in aMT6s secretion as ambient light gradually decreases from morning to evening.
On the other hand, aMT6s levels in samples collected after each stage (evening) significantly decreased throughout the three week period. This suggests an attenuation of exercise induced aMT6s secretion during this period. Similarly, previous research on subjects who were not highly trained showed that the increase in plasma melatonin that occurs after a bout of exercise is attenuated by regular vigorous training.5
MORNING LEVELS OF URINARY AMT6S
In contrast with the acute melatonin response to exercise bouts, morning levels of urinary aMT6s (measured in samples collected between 0900 and 0930, after an overnight rest in darkness) significantly decreased throughout the study. No data are available in the literature on the chronic effects on morning aMT6s levels of such demanding (in terms of both duration and intensity) endurance exercise programmes as the one described here.
Our data suggest a night time reduction in melatonin synthesis induced by physical exercise. The reasons for this phenomenon are not apparent. One could speculate that, with strenuous repeated daily bouts of daytime physical exercise possibly associated with high rates of melatonin secretion, there is not sufficient time overnight for the gland to recover. Concerning the influence of physical exercise performed at night, when pineal melatonin synthesis is already stimulated, previous studies in humans41 have clearly shown that cycling at 50–80% of the personal maximal work capacity significantly blunts the nocturnal increase in plasma melatonin levels. Research with rats42 has shown that swimming at night produced no change in the activities of the enzymes involved in melatonin synthesis and the contents of two melatonin precursors, suggesting that swimming depresses pineal melatonin content by enhancing melatonin efflux from the gland. Catecholamines secreted from the sympathetic nerves, however, are not involved in eliciting the dramatic reduction in elevated pineal melatonin levels.43 Further research is necessary to elucidate the mechanisms involved in the exhaustion that appears to occur in the pineal gland after consecutive bouts of strenuous exercise. Our results indicate for the first time a reduced night time secretory capacity of the epiphysis in parallel with development of the race. The same pattern of exhaustion over the three week period may also have occurred in the adrenal gland and testis, as discussed below.
HPT AND HPA AXES
Testosterone and cortisol levels showed the same pattern during the study—that is, a consistent significant decrease throughout the three week period. It should be emphasised, however, that the mean levels of these two hormones remained within normal physiological limits44 throughout the study. The decrease in resting testosterone found in our subjects is in agreement with previous research, which also reported subclinical decreases in basal testosterone levels of endurance trained humans,15–,19 including competitive cyclists.20 Several pathophysiological mechanisms have been proposed to explain the perturbation of the HPT axis often reported in endurance trained men, and include: (a) loss of body mass and/or energy imbalance associated with strenuous repeated exercise bouts45; (b) altered central stimulation of the gonads—for example, through suppression at the hypothalamic or pituitary level by other hormonal systems such as cortisol or prolactin46; (c) peripheral factors—for example, intrinsic failure of steroid biosynthesis in the testis.46 The first factor can be reasonably ruled out given that the body mass of the subjects remained stable overall during the study. It has, indeed, been previously documented that professional cyclists are able to balance their energy intake with their energy expenditure on a day to day basis, even during three week tour races.37 Given that (a) both FSH and LH remained unchanged during the race and (b) cortisol and aMT6s showed the same overall response as testosterone, the observed testosterone decrease does not seem to be attributable to hormonal mediated suppression of the HPT axis. This hypothesis is partly limited by the fact that prolactin levels were not measured in our study. Thus it remains to be elucidated whether the decrease in testosterone was mediated by the inhibitory effect that training induced hyperprolactinaemia may exert on the HPT axis.46 Concerning the latter, it must be kept in mind that published data are contradictory with regard to the effects of endurance training on basal prolactin levels—that is, this hormone can also decrease or remain unchanged with training.11 Thus our results may be explained by peripheral mechanisms, such as greater use of testosterone by muscle tissue and/or greater hepatic clearance of the hormone47 throughout such a long period of strenuous physical activity. Although further research is necessary, one could expect exhaustion of testosterone production to a certain extent as the result of these peripheral factors. To date, however, training induced failure of gonadal biosynthesis has not been shown in endurance athletes. Similarly, one could also speculate about the possible impact of physical factors on testicular function—for example, increases in intrascrotal temperature and/or testicular microtrauma46 (cyclists must exercise several hours a day wearing tight shorts, during which testis suffer continuous friction against the saddle). However, these physical factors must be viewed with caution, as other glands (epiphysis and adrenal cortex) not affected by these physical factors in our subjects appeared to suffer a certain degree of exhaustion.
A decrease in cortisol was also observed throughout the three week period. Some authors have found the opposite—that is, an increase in basal cortisol levels after long term training13 or in overtrained athletes.14 Seidman et al13 reported a certain state of hypercortisolism in previously untrained men after 18 weeks of strenuous physical training, and Barron et al14 found increased cortisol after a four month period in overtrained marathon runners. The lack of agreement between their findings and our results may be explained by the fact that our investigation was on professional cyclists during a prolonged, highly demanding competition and after an eight month cycling season (during which they had accumulated about 30 000 km and about 90 competition days). To the best of our knowledge, no previous data are available on the endocrinal adaptations to such a demanding endurance sport and after such a long competition season. In line with our findings, Wheeler et al16 found a decrease in basal cortisol levels of endurance runners after six months of training. Bearing in mind the model of both high intensity and long duration of exercise chosen for our investigation, one cannot rule out a certain degree of exhaustion of the adrenal cortex. In fact, a decreased sensitivity of this gland to ACTH stimulation or of the HPA axis to cortisol negative feedback was previously observed by Barron et al14 in overtrained marathon runners after four months, despite increased basal cortisol levels. Similarly, Luger et al48 reported diminished responses of ACTH and cortisol to corticotrophin releasing hormone in highly trained runners with hypercortisolism. One could speculate that a decrease in cortisol concentration caused by adrenal exhaustion would have finally occurred in the subjects of both studies if the training period had been prolonged any further and/or if the training loads had been as demanding as those of professional cycling stage races. Moreover, one can draw an analogy with the decrease in catecholamine levels reported during maximal exercise in overtrained athletes, which is attributable to adrenal exhaustion or the so called parasympathetic form of overtraining.49 Although our study is limited by the fact that we did not measure ACTH, from the findings of studies with highly trained48 or overtrained humans,14 and highly trained rats,50 it could be expected that an inhibitory and/or exhausting mechanism was responsible for the decrease in cortisol found in our subjects—that is, decreased sensitivity of the HPA axis to cortisol negative feedback in a prolonged (three week) stressful situation, and/or decreased responsiveness of cortisol to ACTH stimulation.
Finally, the lack of changes observed in basal FSH and LH is in line with several previous studies.16,18,24 Although some controversy exists, the training induced decrease in basal testosterone often reported in endurance athletes does not seem to be attributable to alterations at the hypothalamic-pituitary level. In studies showing reduction in LH pulse frequency22 and amplitude23 in male endurance athletes, the duration of blood sampling was limited to less than 24 hours (eight and six hours respectively), making it difficult to actually discern the effects of training on the pattern of gonadotrophin secretion.
In conclusion, our data suggest that extreme daytime endurance exercise may increase melatonin secretion, although further studies performed under well controlled conditions are needed to confirm this hypothesis. On the other hand, the activity of the pineal gland, as well as that of other hormonal systems (adrenal glands and testis), may become exhausted after long term exercise. Further research may determine the physiological consequences of this state of hormonal exhaustion on other body systems such as the immune system.
Acknowledgments
This study was financed by Agrupación Deportiva Banesto. It has been awarded the second Premio Nacional de Investigación en Medicina del Deporte 2000 of the Universidad de Oviedo, Spain.