Article Text

Abstract
Objectives: A COL5A1 gene variant was shown to be associated with chronic Achilles tendinopathy in a South African population. The aim of this case–control genetic association study was to investigate the BstUI and DpnII restriction fragment length polymorphisms (RFLP) in a second population from Australia and to identify a predisposing haplotype for Achilles tendinopathy in both populations.
Methods: 85 Australian and 93 South African patients with tendinopathy, as well as 210 Australian and 132 white South African control subjects were genotyped for the BstUI (rs12722) and DpnII (rs13946) RFLP, as well as markers rs10858286, rs3196378, rs11103544, rs4504708 and rs3128575.
Results: The BstUI RFLP (p<0.001) and marker rs3196378 (p = 0.016) were associated with chronic Achilles tendinopathy in Australian subjects. Individuals within both populations with a CC genotype for the BstUI RFLP had a significantly decreased risk of developing tendinopathy versus any other genotypes (Australian odds ratio 0.42, 95% CI 0.20 to 0.86, p = 0.017). The TC inferred haplotype (rs12722, rs3196378) was found to be overrepresented (global p = 0.008) in the South African tendinopathy group compared with all other haplotypes.
Conclusion: The BstUI RFLP is associated with chronic Achilles tendinopathy in a second population and a region within the COL5A1 3′ untranslated region may predispose individuals to an increased risk of developing chronic Achilles tendinopathy.
Statistics from Altmetric.com
An injury to the Achilles tendon is a common clinical presentation that is frequently seen in clinical sports medicine or rheumatological practices.1 The lifetime prevalence of Achilles tendon injury has been reported to be as high as 11%.2 Furthermore, as much as 30–50% of all sporting injuries involve tendons, of which 6–18% comprise Achilles tendon injuries.3 There is a spectrum of pathology that can affect the Achilles tendon and surrounding structures4 most commonly due to overuse (commonly referred to as either “tendinopathy” or “tendinosis”).5
Identification of the different risk factors for Achilles tendon injuries may allow for the design and implementation of successful treatment as well as preventive strategies. The exact aetiology of Achilles tendon injuries, in particular tendinopathy, is, however, not well defined. It is generally accepted that these injuries are complex conditions and that several extrinsic and intrinsic risk factors contribute to the development of Achilles tendinopathy. In recent years, there has been a specific interest in identifying intrinsic risk factors, in particular a genetic predisposition, to Achilles tendon injuries.6 7 The clinical relevance of identifying genetic risk factors associated with Achilles tendon injury is (1) that identification of a genetic risk factor may be of value to ascertain whether repetitive mechanical loading during occupational or sports activity can lead to an increased risk of injury in an individual; (2) perhaps, in future, to assist in determining which therapeutic interventions could be most effective in different individuals with this injury and (3) genetic risk factors may be important in defining the prognosis for this injury.
To date, polymorphisms within two genes, TNC and COL5A1, have been associated with Achilles tendon injuries.8 9 The TNC gene (chromosome 9q33) encodes the tenascin C protein, an important component of tendons; its expression is regulated by mechanical loading.10 11 The COL5A1 (9q34.2–q34.3) gene encodes the proα1(V) chain, the rate-limiting component of type V collagen trimer assembly.12 Type V collagen, belonging to the fibrillar subfamily of collagens, is composed of several isoforms; α1(V)2α2(V) is the most abundant and widely distributed isoform and typically co-assembles with type I collagen to form heterotypic type I/V fibrils in tissues such as tendon, ligament, bone, sclera and cornea.13 14 The type V collagen heterotrimer has been implicated in the regulation of type I collagen fibril diameter; however, the biological functions of this collagen still remain ill defined.15 Although type V collagen is a quantitatively minor fibril-forming collagen, there is evidence to suggest that functionally it represents a major collagen of developing connective tissues.16 For this reason, it is not surprising that several disease-causing mutations have been identified in COL5A1 to cause Ehlers–Danlos syndrome, a heritable connective tissue disorder characterised by laxity and fragility of soft connective tissues such as tendons, ligaments, dermis, sclera, cornea and blood vessels.17 Therefore, it is plausible that sequence variants within the COL5A1 gene, as previously described in a South African population,9 may also predispose individuals to an increased risk of tendon or ligament injuries. It is therefore important to evaluate the reported sequence variant used in the South African study in other independent populations. These studies will provide further evidence highlighting the biological pathways involved in tendon injuries and more specifically the role of type V collagen in Achilles tendon pathology.
The aim of this study was therefore to validate the initial association study of the BstUI restriction fragment length polymorphism (RFLP) (marker rs12722) within the 3′ untranslated region (UTR) of the COL5A1 gene with Achilles tendinopathy in a second white (Australian) population and to identify an associated haplotype for Achilles tendinopathy in both the white South African and white Australian populations.
MATERIALS AND METHODS
Subjects
Eighty-five white patients with documented Achilles tendinopathy (AUS TEN) were recruited by the Musculoskeletal Research Centre at La Trobe University in Melbourne, Australia. One of the investigators (JC) made the diagnosis of chronic Achilles tendinopathy using clinical criteria as previously described.8 9 In all the subjects, soft tissue ultrasound examination was performed to confirm the diagnosis of the affected Achilles tendon.
A total of 210 apparently healthy white control (AUS CON) subjects without any history of any tendon or ligament pathology was also recruited for this study from Melbourne in Australia. The AUS CON subjects were matched for age of initial onset of Achilles tendinopathy in the AUS TEN group. To avoid any possible effects of population stratification, the AUS TEN and AUS CON groups were similarly matched for their country of birth.
Volunteers were required to complete an informed consent form according to the Declaration of Helsinki and once recruited participants detailed personal particulars and medical history by means of a questionnaire. Potential participants who had a history of current or past fluoroquinolone antibiotic use or previous local corticosteroid injections in the Achilles tendon or the area surrounding the Achilles tendon before the onset of symptoms were excluded from the study. This was necessary because of the known association between fluoroquinolone antibiotic18 or possibly corticosteroid use and an increased risk of Achilles tendon rupture.19 Furthermore, potential subjects who had been diagnosed with any connective tissue disorders or any other systemic diseases believed to be associated with Achilles tendon pathology, such as, but not limited to, Ehlers–Danlos syndrome, benign hypermobility joint syndrome, rheumatoid arthritis, systemic lupus erythematosus, hyperparathyroidism, renal insufficiency, diabetes mellitus and familial hypercholesterolaemia were also excluded from the study.19
The subjects for the South African study were recruited as described previously.8 9 In summary, 93 physically active white subjects with a clinical diagnosis of chronic Achilles tendinopathy (SA TEN) and 132 physically active white subjects without any history of clinical symptomatic Achilles tendon injuries (SA CON) were recruited for this study. The SA TEN and SA CON groups were matched for age, gender and to avoid possible effects of population stratification, country of birth.
The study was approved by the Research Ethics Committee of the Faculty of Health Sciences within the University of Cape Town, South Africa (reference numbers 289/2004 and 086/2005) and the Human Ethics Committee of La Trobe University, Melbourne, Australia.
DNA extraction
For the Australian subjects, DNA was extracted from approximately 4.5 ml of venous blood using a sequenced extraction technique (FlexiGene DNA Kit, Qiagen P/L, Valencia, California, USA) as per the manufacturer’s recommendations. DNA for the South African study was isolated as described previously.8 9
SNP analyses
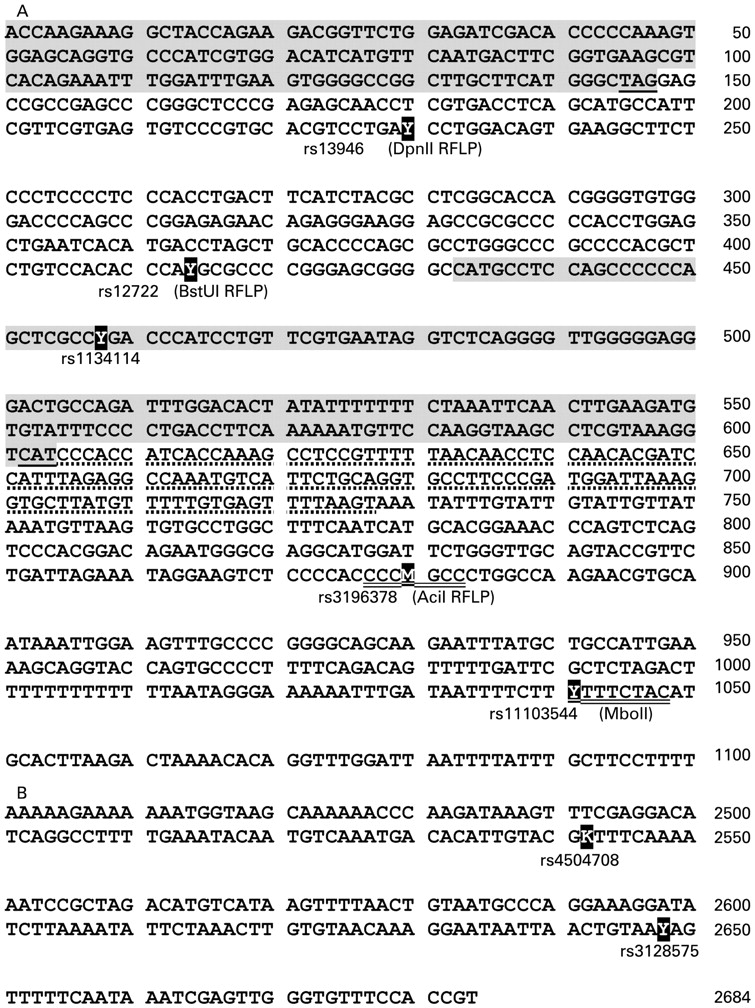
The strongest variant (rs12722, BstUI RFLP) associated with chronic Achilles tendinopathy in the South African study9 was evaluated in a second case–control series of subjects from Australia, as were six other variants (rs10858286, rs13946 (DpnII RFLP also previously used in the South African dataset),9 rs3196378, rs11103544, rs4504708 and rs3128575). Two of the SNP (rs3196378 and rs11103544) were chosen because they were located in putative microRNA binding sites (http://compbio.utmem.edu/miRSNP/ and www.patrocles.org). The relative positions of these variants have been annotated on a schematic representation of the 3′-UTR region of the COL5A1 gene (fig 1) and the genomic sequence of the 3′-UTR region (fig 2A and B).

A schematic representation of the terminal exons (rectangles) and intron (horizontal lines) boundaries of the 3′-end of the COL5A1 gene. The translated regions of the exons are solid whereas the untranslated region (UTR) of exon 66 is clear. The first exon of an expressed sequence tag (EST) (ENSESTG00000033016) identified within the 3′-UTR of COL5A1 is also indicated as a solid rectangle for the translated region and a clear rectangle for the 5′-UTR. The EST is expressed in the opposite orientation to the COL5A1 gene. Exon numbers are indicated and the chromosomal location of the gene is given in brackets. The two exonic single nucleotide polymorphisms (SNP; grey boxes) and the one intronic SNP, rs10858286 (used in this study) identified from databases hosted by the National Centre for Biotechnology Information (NCBI) and the Ensembl Genome Data Centre (Ensembl) are annotated. The accession numbers of the six SNP within the 3′-UTR region of COL5A1 used in this study are also annotated and the base changes are indicated for all the SNP. The minor alleles and the frequencies are also indicated. When applicable the restriction fragment length polymorphism (RFLP) associated with the SNP are indicated. All the information used to construct this figure was obtained from databases hosted by the NCBI (http://www.ncbi.nlm.nih.gov/) and Ensembl http://www.ensembl.org/index.html.

Nucleotide sequence of COL5A1 exon 66 containing (A) the first five sequence variants and (B) the latter two sequence variants investigated in this study. The positions of the single nucleotide polymorphisms (SNP) are annotated in bold (Y = C or T; M = A or C and K = G or T). The accession numbers of the SNP are given, including, when appropriate, the name of the restriction fragment length polymorphism in brackets. The nucleotide position of exon 66 is indicated on the right. The shaded sequences represent the translated region of the COL5A1 exon, 66 (nucleotides 1–147) and exon 1 of the expressed sequence tag (EST) (nucleotides 433–604). The stop codon of exon 66 and the translation start codon of exon 1 is underlined. The dotted line indicates the 5′ untranslated region of the EST, whereas the putative recognition sequences of the two micro RNA are double underlined.
The South African subjects were re-genotyped for variant rs12722. The strongest of the additional variants identified in the Australian subjects (rs3196378) and its flanking marker (rs11103544) were then evaluated in the South African groups. Haplotype frequencies were inferred for both the Australian and South African groups based on the genotype data of variants rs13946, rs12722, rs3196378 and rs11103544.
PCR conditions
The BstUI RFLP was used for the allelic discrimination of variant rs12722, as previously described.9 Three methods were used to discriminate between the alleles of SNP rs3196378, which spans the one miRNA binding site, the AciI RFLP analysis and two primer-specific PCR reactions that amplified each of the two alleles.20 Similarly the MboII RFLP was used for SNP rs11103544, which spans the other miRNA binding site, as previously described.20
Primer pairs for the remaining variants (rs10858286, rs13946, rs4504708 and rs3128575) were designed (table 1). The tetra-primer amplificatory refraction mutation system method21 was used for the allelic discrimination of SNP rs10858286, rs4504708 and rs3128575. PCR amplification was performed in 25–60 μl volumes containing 200 ng genomic DNA; 10–20 pmol of each primer; 2.0–3.0 mmol magnesium chloride/magnesium sulphate, 50 mmol potassium chloride; 10 mmol Tris-HCl (pH 8.3); 200 μmol dATP, dCTP, dGTP and dTTP and 0.5 U Taq DNA polymerase (New England Biolabs, Ipswich, Massachusetts, USA). Amplifications were conducted by denaturing at 95°C for 5 minutes; five cycles of denaturation at 95°C for 25 s, annealing at either 55, 59, 60, 62 or 70°C for 45 s and extension at 72°C for 30 s; 27 cycles of denaturation at 95°C for 25 s, annealing at 50°C for 45 s and extension at 72°C for 30 s and a final extension at 72°C for 10 minutes (XP Thermal Cycler Block; Bioer Technology Co; Middlesex, UK).
The resultant PCR fragments were separated together with a 100 bp DNA size standard (Promega Corporation, Madison, Wisconsin, USA) on 2% agarose gels and visualised by SYBER Gold nucleic acid gel stain (Invitrogen Molecular Probes, Oregon, USA). The gels were photographed under ultraviolet light using a photodocumentation system (Uvitec Ltd, Cambridge, UK) and the sizes of the DNA fragments were determined.
Statistical analyses
Quantitative baseline variables, such as height, between the tendinopathy and control groups, were compared with analysis of variance, adjusting for age and gender when required. Chi-squared tests or Fisher tests, when necessary, were used to compare categorical variables between the diagnostic groups and between countries. We did not adjust for multiple testing. The programming language and environment R (http://www.R-project.org) and STATISTICA version 7 were used for basic descriptive statistics. The R package, genetics, was used for estimating all frequencies, Hardy–Weinberg equilibrium (HWE) probabilities and testing linkage disequilibrium (LD). Haplotype frequencies were inferred and analysed using the R package, haplo.stats. The R package, LDheatmap, was used to construct the LD maps.
RESULTS
Subject characteristics
The AUS TEN and AUS CON groups were similarly matched for age, dominant hand and country of birth (table 2). The age of the AUS TEN subjects refers to the age of the initial onset of Achilles tendon symptoms and they were recruited on average 8.9 years (SD 9.8) after the onset of their initial symptoms. The AUS TEN subjects were on average significantly heavier (p<0.001) with correspondingly higher average body mass index (BMI) (p<0.001) than the AUS CON subjects. Similarly, the AUS TEN subjects also contained significantly more men than the AUS CON subjects (p<0.001). The significant differences in height (p = 0.057) and weight (p = 0.134) between the AUS TEN and AUS CON subjects were no longer significant after adjusting for gender.
Forty-one (48.2%) of the AUS TEN group were diagnosed with unilateral (21 right, 24.7%, and 20 left, 23.5%) chronic Achilles tendinopathy and 44 (51.8%) with bilateral chronic Achilles tendinopathy. Twenty-three (39.0%) subjects reported multiple injuries to the Achilles tendon. Eighty-one of the 85 subjects were symptomatic (95.3%), symptoms were of gradual onset in 62 and more acute in 19 of these subjects.
A total of 112 (94.1%) of the 119 AUS CON subjects whose Achilles tendons were examined by ultrasound were normal. Achilles tendinopathy was confirmed in the four asymptomatic subjects by ultrasound examination and the ultrasound examination was normal in four subjects with symptoms. Five (5.9%) subjects were diagnosed with a partial rupture of the Achilles tendon. The anatomical site of pathology in the subjects was in the middle in 78 subjects (91.8%), distal in five subjects (5.9%) and at the site of insertion in two subjects (2.4%). Thirty-two (37.6%) of the AUS TEN subjects reported a history of injury to other tendons, which included the rotator cuff (N = 12), patellar tendon (N = 1), lateral elbow (N = 13), medial elbow (N = 4) and others (N = 2). None of the tendinopathy group reported corticosteroid injections before the onset of injuries; however, 10 (11.8%) reported having corticosteroid injections after the onset of symptoms.
The South African subjects were previously described in detail.8 9 In summary, the AUS TEN and SA TEN, as well as the AUS CON and SA CON, groups were similarly matched for age of onset, age of recruitment and weight (data not shown). The AUS CON group was, however, significantly shorter (even when covaried for gender) with significantly fewer men than the SA CON group, whereas both the SA TEN and AUS TEN groups were similarly matched for height and gender (data not shown). Finally, both the AUS TEN and AUS CON groups had a significantly higher BMI than their counterpart South African groups (data not shown).
Genotype and allele frequencies
Allele and genotype frequency information for each of the variants is shown in table 3. There was a highly significant difference in the genotype distribution of rs12722 (BstUI RFLP) between the AUS CON and AUS TEN subjects (p<0.001). As previously reported,9 and confirmed in this study, there was a significant difference in the genotype distribution between the SA CON and SA TEN groups (p = 0.024). The genotype distributions of rs12722 were similar between the Australian and South African control groups (p = 0.532), but not the tendinopathy groups (p = 0.036); the AUS TEN group being more heterozygous at this locus than the South African group. However, individuals with a CC genotype had a significantly decreased risk of developing chronic Achilles tendinopathy than those with a T allele (TC or TT genotype) in both the Australian (odds ratio (OR) 0.42, 95% CI 0.20 to 0.86, p = 0.017) and South African (OR 0.38, 95% CI 0.18 to 0.77, P = 0.008) groups.
The differences in the genotype distributions of marker rs13946 (DpnII RFLP) between the control and tendinopathy groups in both the Australian (p = 0.197) and South African cohorts (p = 0.182) were not significant.9 The genotype distributions of both the control (p = 0.436) and tendinopathy (p = 0.646) groups were similar between Australia and South Africa, and as reported for other white populations (http://www.ncbi.nlm.nih.gov/).
A significant difference in the genotype distribution between the AUS CON and AUS TEN groups was noted for rs3196378, which spans a putative miRNA binding site (p<0.016). The AC genotype was significantly overrepresented in the AUS TEN (75%) group when compared with the AUS CON (56%) group (OR 2.3, 95% CI 1.3 to 4.1, p = 0.004). The genotype distribution of this SNP was, however, similar between the SA TEN and SA CON groups (p = 0.464). No significant differences in the genotype distribution between the AUS CON and AUS TEN groups were noted for rs11103544 (p = 0.458), nor between the SA CON and SA TEN groups (p = 0.629), the second marker that spans a putative miRNA binding site.
None of the differences in genotype distribution between the AUS CON and AUS TEN groups for markers rs10858286 (p = 0.468), rs4504708 (p = 0.285) and rs3128575 (p = 0.178) were significant. For this reason, these markers were not analysed in the South African group. The genotype and/or allele frequency distributions of all seven markers presented in this study were similar to previously reported values for white populations (http://www.ncbi.nlm.nih.gov/).
The HWE p values for each polymorphism were calculated and presented in table 3. SNP rs12722 (BstUI RFLP) was not in HWE in any of the Australian groups (p⩽0.012). It was, however, in HWE in both South African groups (p⩾0.080). Except for the SA CON group (p = 0.040), SNP rs13946 (DpnII RFLP) was in HWE in the remaining three groups (p⩾0.477). SNP (rs3196378), which is located within a putative miRNA binding site, was not in HWE for the AUS TEN group (p<0.001) whereas the remainder of the three groups were (p⩾0.063). The remaining SNP (rs10858286, rs4504708 and rs3128575), which were only genotyped in the Australian subjects were in HWE in both the tendinopathy and control groups.
The Australian group showed complete LD for all marker pairs from rs13946, rs12722, rs4504708 and rs3128575 (D′ values were ⩾0.69, independent of diagnostic group; fig 3), thus confirming published LD patterns obtained for other populations. Similarly, marker rs3196378, which overlaps a putative miRNA binding site, was in LD with all of these markers (D′ ⩾ 0.67). A rapid decay in LD was observed for the AUS CON and AUS TEN group between marker rs11103544, which also overlaps a putative miRNA binding site, and rs12722 (D′ ⩽ 0.49). To our knowledge no published LD patterns are available for these two markers. Not surprisingly, marker rs10858286 located within intron 65 of COL5A1 was not in strong LD with any of the markers analysed (fig 3). The LD patterns for the Australian and South African cohorts were similar (fig 3), with the only exception that a rapid decay in LD was noted in the SA TEN group between markers rs1103544 (the second marker spanning a putative miRNA binding site) and rs13946 (D′ = 0.01) and rs13946 and rs12722 (D′ = 0.56). The LD structure obtained for the Australian and South African groups between marker pairs rs13946 and rs11103544 and rs12722 and rs11103544 differed from the published LD structure. Complete LD was noted for these markers in the public databases.

Linkage disequilibrium (LD) structure of the candidate region of the 3′-end of the COL5A1 gene. The LD maps were constructed using genotypes from the Australian control (AUS CON) (left panel) and Australian tendinopathy (AUS TEN) (right panel) groups. The dashed lines delineate the markers genotyped in both Australian and South African subjects. Black and dark grey cells, strong LD; grey cells, intermediate, and light grey and white cells, evidence for historical recombination. The D′ values between pairs of markers are given within each cell. The D′ values for South Africa are given in brackets.
Haplotype analyses
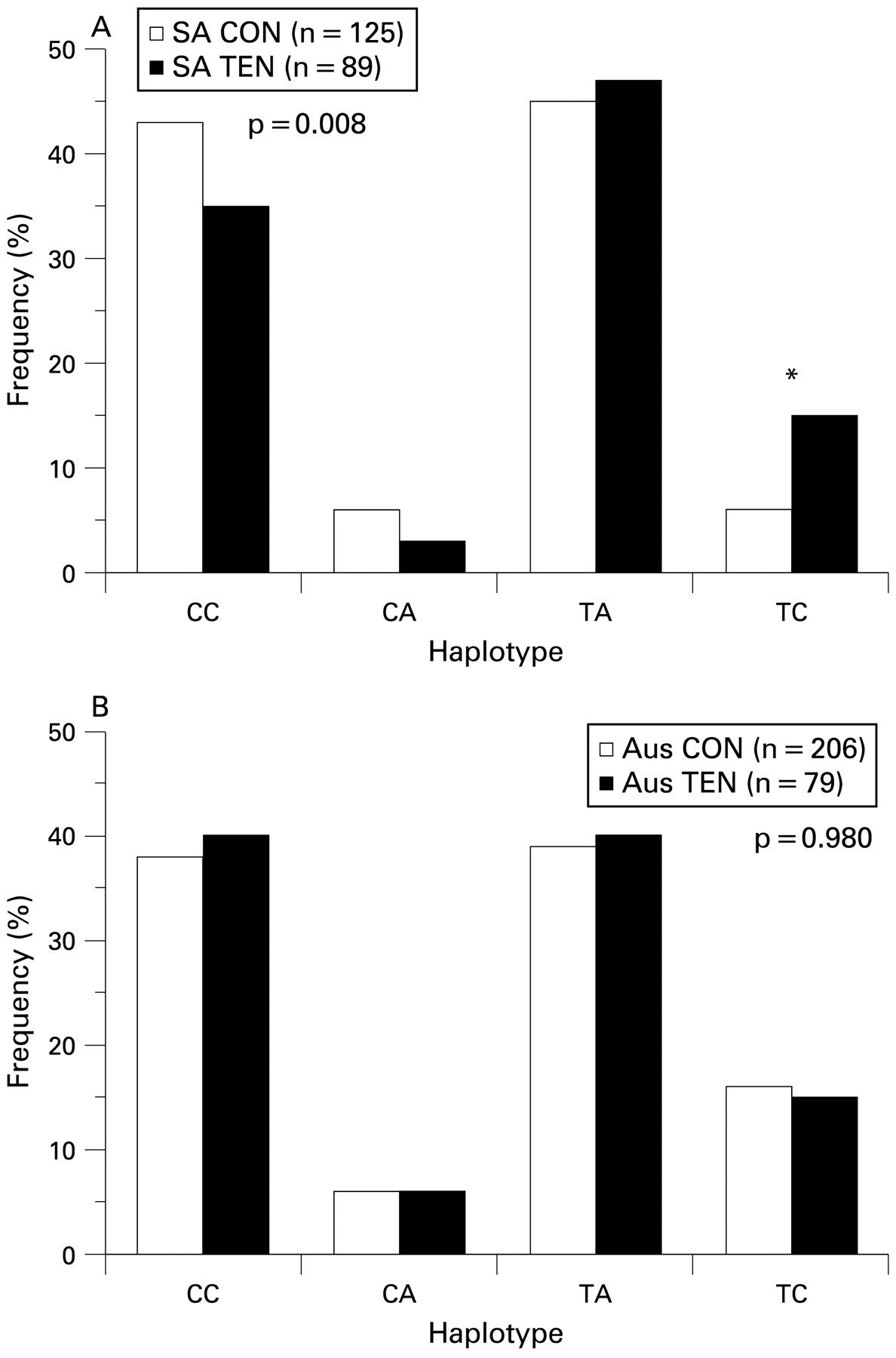
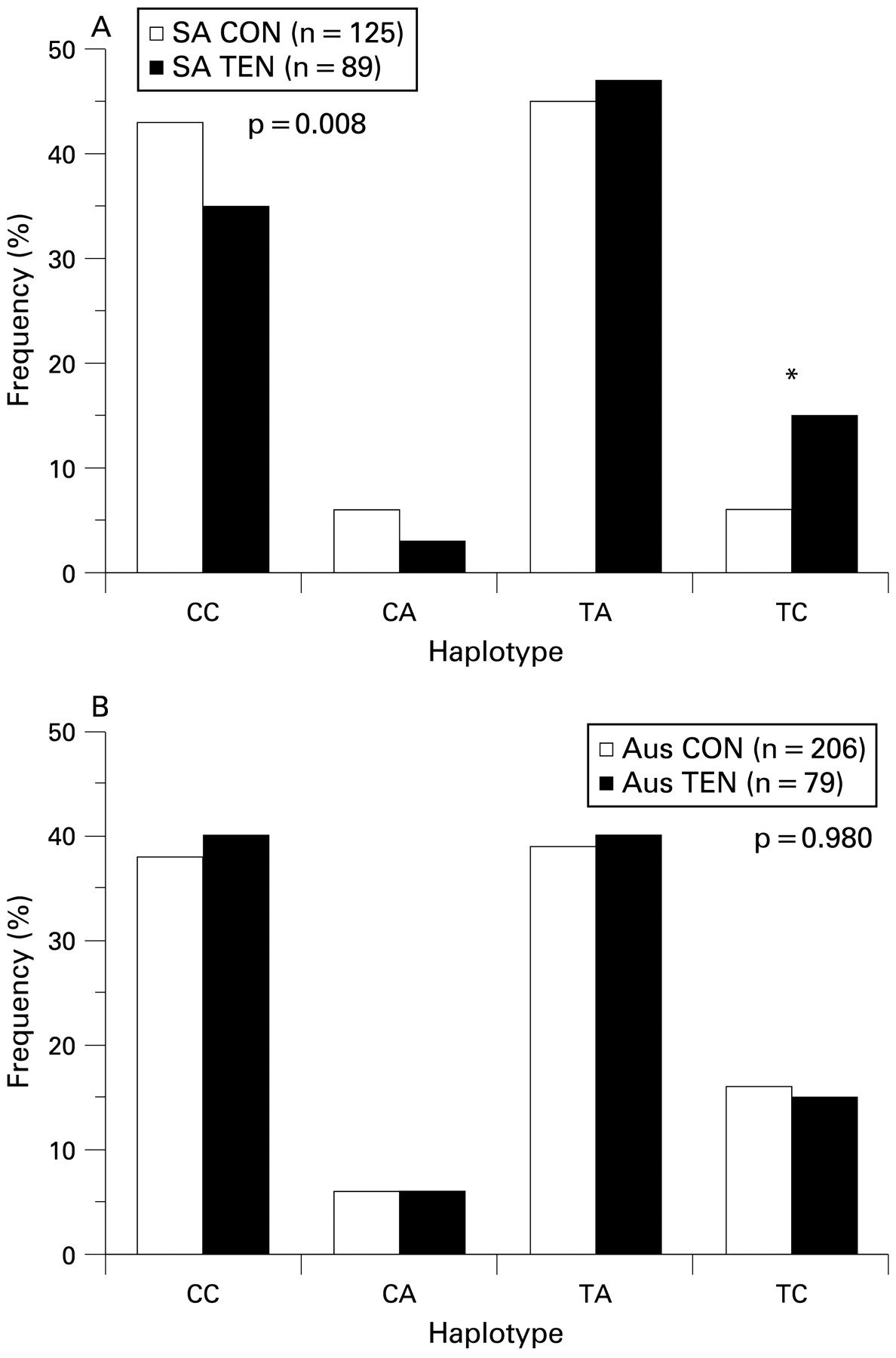
No statistically significant differences in the inferred haplotype (consisting of all four markers) distributions between diagnostic groups were noted in the Australian (global p = 0.280) nor the South African (global p = 0.320) groups (see supplementary table A1, available online only). There was, however, a tendency (p = 0.07) for the TTCT inferred haplotype to be overrepresented in the SA TEN group (7%) when compared with the SA CON group (3%). A significant difference in the inferred haplotype (consisting of markers rs12722 and rs3196378) distribution was, however, observed between the tendinopathy and control groups for South Africa (global p = 0.008). Of note was the overrepresentation of the TC haplotype in the SA TEN (15%) subjects compared with the SA CON (6%) subjects (p<0.001; fig 4A). However, this observation was not mirrored in the Australian group (fig 4B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Inferred haplotype frequency distributions from single nucleotide polymorphisms (SNP) rs12722 (BstUI restriction fragment length polymorphism (RFLP)) and rs3196378 (AciI RFLP) within the 3′ untranslated region (UTR) of the COL5A1 gene in the (A) South African and (B) Australian groups. The clear and solid bars represent the control and Achilles tendinopathy groups, respectively. The global p value for a significant difference in distributions is given, and the asterisk marks the pair with a significant difference in frequency. AUS CON, Australian control group; AUS TEN, Australian tendinopathy group; SA CON, South African control group; SA TEN, South African tendinopathy group.
DISCUSSION
Genetic association studies still remain one of the more common methods used to identify specific regions of the genome that contain genes that predispose individuals to an increased risk of disease, in particular complex conditions such as Achilles tendon injuries.6 7 Achilles tendinopathy is a common injury seen in rheumatological and clinical sports medicine practice, and is associated with repetitive mechanical loading during occupational and sporting activity. Mokone et al9 conducted a case–control association study in a South African population and identified a common allelic variant, marker rs12722 (BstUI RFLP) to be associated with symptomatic chronic Achilles tendinopathy, but not rs13946 (DpnII RFLP); both markers are within the 3′-UTR of the COL5A1 gene. This initial finding has attracted interest from clinicians and basic scientists working in the field of tendon injury.1 However, it is important that the findings of this single genetic association study are replicated in other populations. This is pivotal in providing added strength that any associations identified reflect biological processes that are important in understanding the pathophysiology of the injury.22
This study has shown that there was a highly significant (p<0.001) genotype association at the rs12722 (BstUI RFLP) locus, but not the rs13946 (DpnII RFLP) locus, with symptomatic chronic Achilles tendinopathy in Australian subjects of white ancestry. In addition, this study has shown that individuals with a CC genotype for marker rs12722 had a significantly decreased risk of developing chronic Achilles tendinopathy compared with those with a T allele (TC or TT genotype) in both the Australian (OR 0.42, 95% CI 0.20 to 0.86, p = 0.017) and South African (OR 0.38, 95% CI 0.18 to 0.77, p = 0.008) subjects. The functional significance of this polymorphism is to our knowledge still unknown. This association therefore provides further evidence supporting the hypothesis that sequence variant(s) within the COL5A1 gene may predispose individuals to an increased risk of chronic Achilles tendinopathy.
In an attempt to identify a predisposing haplotype in both the Australian and South African groups, additional markers were selected for genotyping. The differences in the genotype or allele distribution of the upstream marker rs10858286, within intron 65, and the markers rs4504708 and rs3128575 located within the 3′-UTR, 2129 and 2135 bp downstream of rs12722, respectively, were not significant for the Australian group and therefore they were not evaluated in the South African individuals.
Additional markers rs3196378 and rs11103544 localised to the 3′-UTR, 466 and 628 bp downstream of rs12722 (BstUI RFLP), were selected because it was predicted that they span putative miRNA recognition sequences, CCCCGCC and TTTTCTAC (polymorphic nucleotides underlined), respectively, and therefore may potentially have a functional significance (http://compbio.utmem.edu/miRSNP/ and www.patrocles.org). A significant association at the rs3196378 locus was noted in the Australian group (p = 0.016) only. Individuals with the heterozygous TC genotype had a significantly increased risk of developing chronic Achilles tendinopathy compared with those with a homozygous TT or CC genotype. Interestingly, a higher heterozygous frequency was also noted in the Australian group at the BstUI RFLP locus, which was in LD with marker rs3196378. The reason for this observation in the Australian subjects is not apparently obvious; however, this phenomenon of increased heterozygosity (referred to as heterosis) and disease risk has previously been observed.23 These associations nevertheless provide further evidence supporting the hypothesis that sequence variant(s) within the 3′-end of the COL5A1 gene may predispose individuals to an increased risk of chronic Achilles tendinopathy.
The frequency distribution of sequence variants including SNP can vary between different populations and, in addition, the LD structure can also differ between populations.24 Both these observations have been noted in this study and therefore inferred haplotype frequencies were also investigated because they can provide more information about a segment/sequence region on a chromosome compared with a specific single base sequence.25 26
A departure from HWE was noted for the markers rs13946, rs12722 and rs3196378 in some of the groups. It should be noted that these three markers are within the region of the 3′-UTR associated with tendinopathy. This observation suggests that perhaps the individuals investigated (controls and affected subjects) in this study do not represent a randomly mating population or the sample size of this study is too small.27 The controls investigated were selected based on clinical diagnosis, age, geographical ancestry and physical activity to match the affected subjects, who were selected based on clinical criteria and geographical ancestry. For this reason, the individuals included in this study were highly selective and this could therefore be the more likely reason why a departure from HWE was noted for these markers. It is evident that more population data for all three of these markers need to be accumulated to re-evaluate the HWE values on a larger sample size. The flanking markers (rs10858286, rs11103544, rs4504708 and rs3128575) were, however, in HWE in both the control and tendinopathy groups of the Australian and South African cohorts.
The haplotype analyses using combinations of markers rs13946, rs12722, rs3196378 and rs11103544 for the Australian and South African groups revealed significant differences in the frequency distributions between the control and tendinopathy groups of the South African study. Of particular interest was the significant overrepresentation (p<0.001) of the TC haplotype (consisting of markers rs12722 and rs3196378) in the SA TEN group.
What this study adds
The COL5A1 BstUI RFLP is associated with Achilles tendinopathy in two independent populations. The results from this study implicate a critical interval within the 3′-UTR COL5A1 with an increased risk of Achilles tendinopathy.
It is interesting to note that the TC haplotype observed more frequently with the tendinopathy group includes the sequence variants (underlined) for the miRNA recognition sequences at marker rs3196378. miRNA are a class of small non-coding RNA that function as key regulators of gene expression and are able to regulate the expression of genes at a posttranscriptional level by specific inhibition of translation or by inducing mRNA cleavage.28–31 This miRNA-mediated gene silencing is a complex process that involves the binding and interactions of different proteins, including the potential binding of several miRNA to achieve the desired expression level of the target protein. Sequence variations including SNP within the miRNA target sequence and other critical sequence regions required for posttranscriptional regulation have the potential to affect miRNA binding, which may lead to significant modification in protein expression and phenotypic/clinical consequences.32
We hypothesise that individuals who inherit the haplotype containing the altered miRNA recognition site may result in the altered binding of the specific miRNA and translation of COL5A1. This may lead to unfavourable levels of type V collagen expression, which may subsequently affect the type V : type I collagen ratios, leading to a compromised collagen fibre. As previously described, type V collagen expression levels are critical in determining fibre diameter and strength.13 14
The association of the individual markers rs12722 and rs1103544 in the Australian group, together with the common inferred haplotype of the same two markers among the SA TEN group, implicates the critical interval containing the predisposing allele to a region within the 3′-UTR of the COL5A1 gene. It is now necessary to evaluate these markers and haplotypes further in other populations so that more evidence can be accumulated to support the proposed hypothesis presented above. The functional significance of markers rs12722 and rs3196378 also needs to be determined in relation to type V collagen expression levels or activity. It would be of further interest to evaluate the significance of these particular markers and common haplotypes in other soft tissue injuries, the repair process and defining the prognoses of these injuries.
In conclusion, this study has shown that the BstUI RFLP (rs12722) is associated with chronic Achilles tendinopathy in a second population as tested in Australian subjects of white ancestry. In addition, this study has identified a critical region of the 3′-UTR of the COL5A1 gene containing the potential causal sequence that may predispose individuals to an increased risk of developing chronic Achilles tendinopathy. Although we recognise that chronic Achilles tendinopathy is a complex condition that is caused by the interactions of several genetic and non-genetic risk factors, this case–control investigation has identified a critical interval within the COL5A1 3′-UTR that predisposes individuals to develop chronic Achilles tendinopathy in subjects from both Australia and South Africa. Clinicians treating patients with chronic Achilles tendinopathy should be made aware of the evidence that genetic factors may, partly, play a role in determining the risk of developing this injury. Furthermore, the possible role that genetic factors may play in determining responses to therapeutic interventions and therefore affecting the prognosis of chronic Achilles tendinopathy requires further investigation.
Acknowledgments
The authors would like to thank Dr GG Mokone for assistance in the recruitment of the subjects and the phenotypic data collection for the South African Cohort.
REFERENCES
Footnotes
Competing interests: None.
Funding: This study was supported in part by funds from the National Research Foundation (NRF) of South African (grant no FA2005021700015), University of Cape Town and the South African Medical Research Council. AVS was supported by the postdoctoral innovation award from the NRF.
Ethics approval: The study was approved by the Research Ethics Committee of the Faculty of Health Sciences within the University of Cape Town, South Africa (reference numbers 289/2004 and 086/2005) and the Human Ethics Committee of La Trobe University, Melbourne, Australia.
Patient consent: Obtained.
▸ Supplemental table 1A is published online only at http://bjsm.bmj.com/content/vol43/issue5