Article Text

Abstract
It remains unclear by which mechanism ‘live high–train low’ (LHTL) altitude training increases exercise performance. Haematological and skeletal muscle adaptations have both been proposed. To test the hypotheses that (i) LHTL improves maximal oxygen uptake (VO2max) and (ii) this improvement is related to hypoxia-induced increases in total haemoglobin mass (Hbmass) and not to improved maximal oxidative capacity of skeletal muscle, we determined VO2max before LHTL and after LHTL, before and after the altitude-induced increases in Hbmass (measured by carbon-monoxide rebreathing) had been abolished by isovolumic haemodilution. We obtained skeletal muscle biopsies to quantify mitochondrial oxidative capacity and efficiency. Sixteen endurance-trained athletes were assigned (double-blinded, placebo controlled) to ≥16 h/day over 4 weeks to normoxia (placebo, n=6) or normobaric hypoxia equivalent to 3000 m altitude (LHTL, n=10). Four-week LHTL did not increase VO2max, irrespective of treatment (LHTL: 1.5%; placebo: 2.0%). Hbmass was slightly increased (4.6%) in 5 (of 10) LHTL subjects but this was not accompanied by a concurrent increase in VO2max. In the subjects demonstrating an increase in Hbmass, isovolumic haemodilution elicited a 5.8% decrease in VO2max. Cycling efficiency was altered neither with time nor by LHTL. Neither maximal capacity of oxidative phosphorylation nor mitochondrial efficiency was modified by time or LHTL. The present results suggest that LHTL has no positive effect on VO2max in endurance-trained athletes because (i) muscle maximal oxidative capacity is not improved following LHTL and (ii) erythrocyte volume expansion after LHTL, if any, is too small to alter O2 transport.
Statistics from Altmetric.com
Introduction
The concept of ‘live high–train low’ (LHTL) was introduced to preserve the ability to train at a high intensity while acclimatising to hypoxia1 and has repeatedly demonstrated a positive effect on VO2max and sea-level performance.2 ,3 Although the ergogenic effect of LHTL has not been unequivocally confirmed, LHTL is nonetheless considered a strategy to increase aerobic performance,4–6 even if the potential improvement remains modest.7
Assuming that there is an effect of hypoxic exposure on VO2max, the question arises which physiological mechanism(s) might be involved. Two hypoxia-related mechanisms were proposed, one being ‘central’, that is, an altitude-induced increase in total red blood cell volume (RCV)4 and the other mechanism being an increase in skeletal muscle efficiency.5 The rationale for an increased RCV (or total haemoglobin mass, Hbmass) to be the main mechanism of LHTL is that (i) RCV is increased in proportion to the residing altitude,8 (ii) when sufficient altitude ‘time’ is allowed, RCV is increased in most athletes participating in LHTL studies,2 ,9–15 (iii) augmentation of RCV with autologous blood transfusion16 or recombinant erythropoietin17 ,18 increases aerobic performance and (iv) in those athletes for whom RCV is increased in response to LHTL, also VO2max is increased.19
However, it is argued that attributing performance increases solely to enhanced erythropoiesis and RCV is inadequate,5 since some LHTL studies have observed only minor change in RCV whereas at the same time running economy was improved,20–22 suggesting increased muscular efficiency. Although the underlying mechanism has not been demonstrated, it may be that acclimatisation to altitude could improve oxidative enzyme activity and/or mitochondrial function, as shown after a hypoxic training intervention (live low–train high).23 The suggestion of an effect of hypoxia on training-induced muscle adaptation is, however, not supported by muscle enzyme activity24 or exercise economy data.25
Our study aimed to test two hypotheses: (i) normobaric LHTL improves VO2max in endurance-trained athletes, and (ii) improvement in VO2max following LHTL is related to hypoxia-induced increase in RCV (or Hbmass) and not to increased maximal oxidative capacity of skeletal muscle. Our previous report indicates not only that LHTL did not improve VO2max in our group of 10 athletes (hypothesis 1 was therefore not verified) but also that Hbmass response to LHTL was variable, being positive in only half of the LHTL subjects.26 The present paper examines the role of increased Hbmass on VO2max response to LHTL (hypothesis 2) by means of isovolumic haemodilution and reports on skeletal muscle mitochondrial function to gain insight into the mechanisms that may be responsible for the potential change in cycling efficiency.
Methods
The present experiment is part of a double-blind, placebo-controlled study on LHTL reported in detail elsewhere.26
Sixteen (15 males and 1 female) trained cyclists (n=13) and triathletes (n=3), age 29±6 years (mean±SD), height 179±8 cm and body weight 69±9 kg participated in the study after giving oral and written informed consent. The study was approved by the Ethics Committee of Zurich (2010-066/0) and Vaud (215/10) (Switzerland), and conformed to the Declaration of Helsinki.
Study design
All subjects resided at the Centre National de Ski Nordique (1135 m, Prémanon, France) while all experimental procedures were performed at the neighbouring La Vallée hospital (1020 m, Le Sentier, Switzerland).
During the first 2 weeks (lead-in period) all subjects were exposed to the normal environment at Prémanon. For the following 4 weeks (intervention period), the subjects were assigned in a double-blinded, placebo-controlled manner to ≥16 h/day of either normobaric normoxia (placebo group, n=6) or normobaric hypoxia equivalent to 3000 m of altitude (LHTL group, n=10). Two testing sessions were scheduled for the present study, the first one during the lead-in period (baseline), and the second one during the last 3 days of the LHTL intervention period (W4).
Training
Details of the training protocol and results are reported elsewhere.26 The subjects followed their own training habits and were instructed to keep the training intensity and load as stable as possible during the 6 weeks. Training was evaluated daily with a heart rate monitor, and in addition some of the subjects recorded their training load with power metres.27 The average exercise intensity spent (time/week) at a given exercise intensity indicated equivalent training between groups. During weeks W3 and W4, the subjects were asked to taper in preparation for the test sessions. No differences were observed across time and study group with respect to exercise intensity, whereas the quantified total training time decreased significantly with regard to time in both groups but with no difference between the two groups.
Experimental protocols
Incremental exercise to exhaustion was carried out on an electronically braked cycle-ergometer (Monark E-839, Varberg, Sweden). The exercise protocol started with a warm-up period of 5 min at a workload of 150 W followed by 5 min at 200 W (100 and 150 W for the female athlete). Thereafter, the workload was increased by 25 W every minute until exhaustion. Attainment of maximal oxygen uptake was established by standard criteria.28 Expired gas were continuously measured (Quark, Cosmed, Rome, Italy). Breath-by-breath values were averaged over 30 s and the highest averaged value was termed as VO2max. Each subject performed two maximal exercise tests during the lead-in period. The largest VO2max result between the two tests was used to express the baseline VO2max.
Hbmass was quantified by a modified version29 of a carbon monoxide (CO)-rebreathing technique.30 A detailed description of this procedure is given elsewhere.26 Hbmass was determined twice during the lead-in period (baseline) and twice during the fourth week of LHTL intervention (W4). Baseline and W4 values therefore correspond to the average of duplicate measurement. The coefficient of variation for Hbmass, assessed from duplicate baseline during the lead-in period, and expressed as the percent typical error (ie, SD of difference scores/√2), was 2.6%.
Isovolumic haemodilution
To test the hypothesis that augmented RCV is the primary mediator of enhanced VO2max following LHTL, maximal exercise was performed before and after isovolumic haemodilution at the end of the intervention period (W4). We withdrew the amount of extra red blood cells that was gained after LHTL by phlebotomy, and this volume was replaced by 6% hydroxyethyl starch (Voluven 6%, Fresenius Kabi, Bad Homburg, Germany) needed to achieve blood volume and RCV values similar to those measured at baseline. For each subject, the volume of blood to be withdrawn was calculated as ΔHbmass/[Hb]×100, with ΔHbmass being the change in Hbmass with LHTL intervention and [Hb], resting haemoglobin concentration at W4 (measured by a Radiometer ABL800 FLEX analyser). Taking into account analytical variability for Hbmass determination, phlebotomy was conducted only if the volume of blood to be removed exceeded 190 ml. Within 15 min after completion of the first exercise bout, blood was withdrawn into a blood bag (MRG6282L or MCG2272L, Macopharma, Mouvaux, France) positioned on a mixing scale (Docon, Möller Medical, Fulda, Germany), and then stored at 4°C. Hydroxyethyl starch infusion was initiated immediately after phlebotomy. In subjects for whom plasma volume decreased without a concomitant change in Hbmass, only the starch infusion was performed. The subjects were blinded towards the procedure by placing their arm through a screen. In subjects for whom no manipulations were necessary, blood was withdrawn and reinfused to simulate these procedures. The second maximal test was initiated 2 h after the end of the first bout of exercise. After completion of the second exercise test, the subjects were either reinfused with their own blood, or infused with saline, again by using a blinded design. Blood sampling and infusion were performed via an 18-G catheter inserted in an arm vein.
Haematological measurements
Blood viscosity (in centipoises, cps) was measured within 3 h after blood collection in EDTA tubes with a cone/plate viscometer (Model DV-II, Brookfield Engineering Laboratories, Middleboro, Massachusetts, USA) with CPE40 spindle at three shear rates (45, 90 and 225/s).
Skeletal muscle biopsy and preparation of muscle fibres
Under local anaesthetics using the Bergström technique with suction, skeletal muscle biopsies (∼20 mg) were obtained from the vastus lateralis muscle at baseline and at W4. The biopsy was dissected free of fat and connective tissue and thereafter measured for mitochondrial respiration, as previously reported.31 Measurements of oxygen consumption were performed using the high-resolution Oroboros Oxygraph-2k (Oroboros, Innsbruck, Austria). Two substrate-uncoupler-inhibitor-titration protocols were used in this study (i) to measure maximal capacity of oxidative phosphorylation and (ii) to identify specific flux control ratios suggestive of mitochondrial efficiency or coupling capacity (respiratory control ratio and leak control coupling). Analysis of citrate synthase activity is also reported in our previous report.31
Statistics
Nonparametric Mann-Whitney U-test was used to test the effect of LHTL (versus placebo). Within a given group (placebo or LHTL), the effect of time or isovolumic haemodilution (baseline, W4, W4 after isovolumic haemodilution) on the different parameters was evaluated with the use of Friedman non-parametric test and planned pair-wise-specific comparisons were made using the Wilcoxon test. The same procedure was also applied by considering three groups rather than two (ie, placebo (n=6); LHTL(+) (n=5) and LHTL(−) (n=5)), LHTL(+) and LHTL(−), respectively, referring to the subgroups of ‘responders’ and ‘non-responders’, that is, the athletes who did (or did not) increase their Hbmass following 4 weeks of LHTL (W4). Statistics were carried out with the Statview 5.0 software (SAS Institute, Cary, North Carolina, USA). The values are reported as arithmetic means±SD unless otherwise stated. Differences were considered as significant for p<0.05.
Results
Four weeks of normobaric LHTL did not increase VO2max
LHTL resulted in a non-significant increase in VO2max (relative to body mass), being 1.5% in LHTL (n=10) and 2.0% in placebo subjects (n=6) (table 1). Maximal workload was non-significantly augmented by 2.2% in LHTL and by 2.7% in placebo subjects.26
Submaximal exercise, maximal exercise and total haemoglobin mass
LHTL induced an increase in Hbmass in 50% of the athletes
The increase in Hbmass among the 10 athletes exposed to LHTL was marginal since five subjects did not increase Hbmass (LHTL(−)), while the other five experienced a 4.6% increase in Hbmass (LHTL(+)) (table 1 and figure 1). All subjects developed haemoconcentration during the study.26 Blood viscosity was similar between groups, and unchanged after LHTL or placebo intervention (W4).
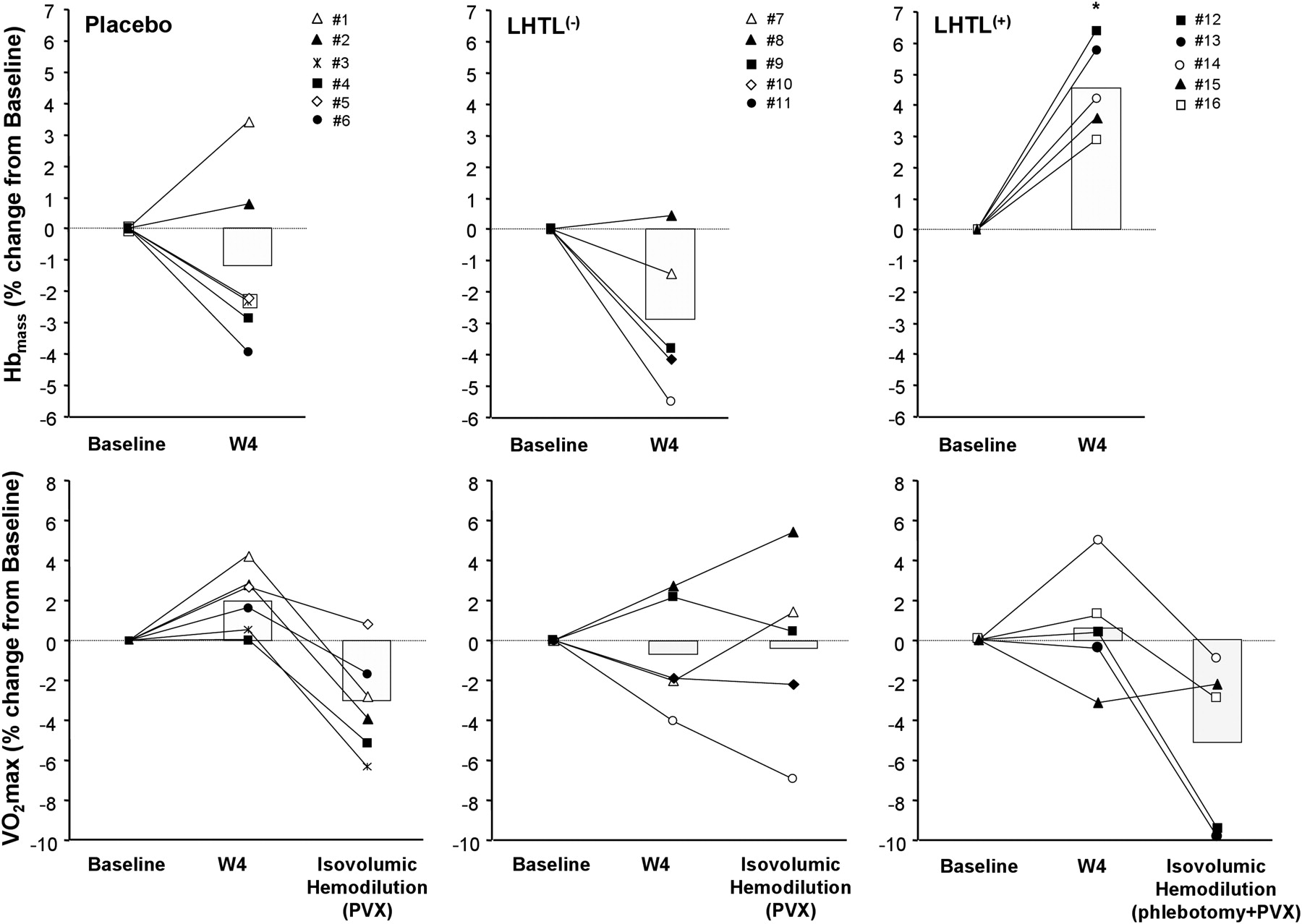
The effects of live high–train low (LHTL) and isovolumic haemodilution on haemoglobin mass (Hbmass) and VO2max. The upper panels show the individual response of total Hbmass (in absolute values), induced by 4 weeks of placebo (n=6) or live high—train low intervention (LHTL; n=10). LHTL athletes were divided, according to their individual changes in Hbmass, into ‘responders’ (LHTL(+), n=5) or ‘non-responders’ (LHTL(−), n=5). Of note is that intervention was associated with a decrease in plasma volume in all subjects.26 The lower panels show first the concurrent individual changes in VO2max (in absolute values), in response to LHTL or placebo intervention (W4 vs baseline), and second the modulation of these changes in response to acute isovolumic haemodilution, aimed at restoring intravascular volumes to basal values. The subjects who gained Hbmass after intervention (ie, LHTL(+)) were isovolumically haemodiluted by means of phlebotomy and plasma volume expander (PVX) infusion, whereas those who did not increase their Hbmass (ie, LHTL(−) and placebo) received PVX infusion only. Histograms indicate means. *p<0.05 vs baseline; **p<0.05 vs W4.
LHTL-induced gain in Hbmass is not associated with a concurrent increase in VO2max
Maximal workload increased in LHTL(+) but not in LHTL(−), suggesting a link between Hbmass and performance gain after LHTL (table 1); however this was not supported by VO2max data, indicating that LHTL(+) athletes experienced a non-significant 1.0% gain in VO2max after LHTL, comparable to the also non-significant 2.1% gain observed among LHTL(−) athletes (table 1 and figure 1).
Suppression of the LHTL-induced gain in Hbmass resulted in a significant reduction of VO2max
LHTL(+) subjects presenting with Hbmass expansion and plasma volume contraction after LHTL underwent blood withdrawal (280±74 ml), followed by plasma expander infusion (540±89 ml), while LHTL(−) and placebo subjects, presenting with no change in Hbmass but plasma volume contraction, received plasma expander only, respectively, 572±299 and 567±169 ml. These interventions allowed for [Hb] to decrease, on average (n=16), from 15.0±0.8 g/dl before isovolumic haemodilution to 14.1±0.7 g/dl after isovolumic haemodilution, that is, at a level comparable to baseline value (14.0±0.5 g/dl).
Isovolumic haemodilution decreased VO2max in LHTL(+), while plasma expander infusion per se had no effect on VO2max in LHTL(−) (table 1 and figure 1). Maximal workload was reduced in LHTL(+) but not in LHTL(−). Plasma expander infusion did not significantly alter maximal workload in placebo athletes but did decrease their VO2max significantly.
Cycling efficiency was not altered following LHTL
Cycling efficiency at 150 and 200 W remained unchanged throughout the study, in both the placebo and LHTL groups.26 Isovolumic haemodilution did not alter cycling efficiency (table 1).
LHTL was not associated with changes in mitochondrial content or function
Mitochondrial content (ie, citrate synthase activity) was altered neither by LHTL nor by time (figure 2A). Mitochondrial function (oxygen flux per unit of mitochondrial content) was similar in both groups, both at baseline and at W4 (figure 2B). The respiratory control ratio and leak control coupling, both indicative of mitochondrial efficiency, were found similar between groups at baseline and unchanged following LHTL or placebo intervention (figure 2C,D).

{kind=link}
{kind=link}
Live high–train low (LHTL) does not alter mitochondrial function of skeletal muscle. The figure shows the mitochondrial function of skeletal muscle in subjects assigned to the placebo (n=6) and LHTL (n=9) groups. Open bars represent baseline measurements and grey bars represent measurements following 4 weeks of respective treatment (W4). Panel A shows citrate synthase activity. Panel B shows maximal oxidative phosphorylation capacity, expressed per unit of citrate synthase activity. Panel C shows respiratory control ratio, presented as the ratio of maximal state 3 respiration through complex I to malate-induced (2 mM) and pyruvate-induced (5 mM) leak respiration. Panel D shows leak coupling control presented as the ratio of malate-induced (2 mM) and pyruvate-induced (5 mM) leak respiration to maximal state 3 respiration through complex I (PI). Values are reported as mean±SE.
Discussion
Using a double-blinded, placebo-controlled design we found that LHTL induced no positive effect on VO2max in endurance athletes. Only 5 of 10 LHTL athletes responded by increasing Hbmass. These ‘responders’ also increased maximal workload but not VO2max in comparison with either their non-responding LHTL counterparts or the placebo athletes. The importance of Hbmass and/or [Hb] for O2 transport during maximal exercise was however confirmed since isovolumic haemodilution resulted in a decrease in both VO2max and maximal workload. Finally, LHTL did not affect skeletal muscle mitochondrial function or whole body cycling efficiency.
Response of Hbmass to LHTL and its role on VO2max
Data analysis revealed that 5 of 10 LHTL subjects did increase Hbmass, suggesting a significant variability among athletes with regard to the haematological response to hypoxia, and that moderate hypoxia may provide, at least in some athletes, a sufficient stimulus for red blood cell synthesis to be increased. Similarly, Chapman et al19 found that only 53% (17 of 32 subjects) exhibited an expansion of RCV in response to altitude training. The different Hbmass response among our LHTL subjects was not explained by the degree of haemoconcentration or by the increase of reticulocyte count. Also an increase in Hbmass following LHTL was not accompanied by a concomitant rise in VO2max, confirming previous data,9 which questions the significance of the small Hbmass improvements detected in athletes undergoing the LHTL regimen. Of note, after blood transfusion, only 50% of the added O2 delivery may be utilised by the tissues.32 Nevertheless, it could be that highly trained individuals require a larger increase in Hbmass to increase VO2max as normal healthy individuals.
In spite of their ‘insensitivity’ to slightly increased O2 delivery, the LHTL athletes were critically dependent on convective O2 transport, since acute normalisation of Hbmass via isovolumic haemodilution provoked a significant drop in maximal power output and VO2max. Similarly, lowering of Hbmass through blood donation33 or haemodilution16 decreases VO2max. On the contrary, the effect of clamping Hbmass during LHTL (via repeated phlebotomy) on maximal aerobic performance was found to be marginal,10 but such procedure substantially differs from acute isovolumic haemodilution.
An important factor to consider during blood withdrawal and/or haemodilution (secondary to plasma expander infusion) is the acute reduction in [Hb], which is known to decrease VO2max, because of a reduction of the oxygen-carrying capacity of blood.34 Although not in line with previous data in athletes indicating no change in VO2max following plasma volume expansion,35 our finding demonstrating that VO2max decreased after haemodilution in the placebo group confirms the important role of [Hb] on VO2max, further suggesting that Hbmass per se is not the only factor controlling VO2max after isovolumic haemodilution in the LHTL(+) group. Conversely, the finding that haemodilution did not affect VO2max in the LHTL(−) group suggests only a minor contribution of [Hb]. Whatever the exact role of [Hb] on VO2max, our data imply that LHTL(−) athletes, unlike placebo subjects, compensated for reduced arterial O2 content by achieving higher maximal cardiac output and/or O2 extraction. Although stroke volume was not measured in the present study, we speculate that cardiac response after isovolumic haemodilution was not much different between placebo and LHTL subjects since (i) infused volume was identical and (ii) decreases in maximal heart rate were small (3–4 beats/min) and similar between groups.
We cannot exclude that incomplete recovery after the first VO2max test may have contributed to some fatigue during the second exercise bout. However, VO2max should have recovered within 2 h36 ,37 and athletes are accustomed to perform repeated bouts of high-intensity exercise.
Mitochondrial function is not improved following LHTL
Hypoxic regulation of mitochondrial expression is not well understood. Skeletal muscle mitochondrial volume density,38 enzyme activities39 and protein content40 have all been observed to decrease following extended hypoxic exposure while other studies have shown negligible differences in mitochondrial-specific biochemical expression.41–44 We have previously found no significant alteration in respiratory chain function in lowland natives measured at sea level and again following 7–9 days of high-altitude exposure,45 and the findings in the current study support these data as mitochondrial function in the LHTL subjects did not differ from data obtained in the control subjects. The present observation at the tissue level provides a basis for explaining why cycling efficiency remained unchanged following LHTL. Other explanations may be related to unchanged motor control and/or biomechanics, which are also determinants of exercise efficiency.46
Conclusion
In the present study VO2max did not increase in response to LHTL and potential mechanisms that could influence VO2max were not identified: we evaluated both central and peripheral adaptations to LHTL and found no effect in highly trained individuals. Furthermore, even though LHTL caused modest erythrocyte volume expansion in some individuals, these subjects did not enhance their VO2max. In conclusion, the positive effects of LHTL on O2 transport appear to be negligible among elite cyclists who already possess very high aerobic capacities conferred by high Hbmass and VO2max.
What this study adds
-
This study provides a mechanistic basis to explain why normobaric live high–train low (LHTL) does not boost VO2max in endurance-trained athletes:
-
The modest increase in haemoglobin mass conferred by LHTL is not sufficient to further increase the athlete's high VO2max, even if VO2max critically depends on Hbmass in this population.
-
Unchanged maximal oxidative capacity or mitochondrial efficiency after LHTL provides evidence against the hypothesis that LHTL may improve muscle efficiency and therefore VO2max.
-
-
Extrapolation to natural altitude is limited since hypobaric hypoxia could induce different responses than those occurring with normobaric hypoxia,47 ,48 although this seems unlikely to have major implications in the present setting.
Acknowledgments
We are grateful to the participants in the study. The authors would like to acknowledge Oroboros Instruments (Innsbruck, Austria) for making valuable equipment at our disposal during the study period. We also would like to thank Ms. Anne Schaerrer and her team (Le Sentier hospital) for excellent logistical support throughout the experiment, as well as the CNSN team (Prémanon) for valuable help during the study.
References
Footnotes
Funding This study was funded through grants obtained from the Bundes Amt für Sport (BASPO, Switzerland), Team Danmark (Denmark) and Ministère des Sports/Institut National du Sport, de l'Expertise et de la Performance (France).
Ethics approval Ethics Committee of Zurich (2010-066/0) and Vaud (215/10) (Switzerland).
Provenance and peer review Not commissioned; externally peer reviewed.