Article Text

Abstract
Administration of recombinant human erythropoietin (rHumanEPO) improves sporting performance and hence is frequently subject to abuse by athletes, although rHumanEPO is prohibited by the WADA. Approaches to detect rHumanEPO doping have improved significantly in recent years but remain imperfect. A new transcriptomic-based longitudinal screening approach is being developed that has the potential to improve the analytical performance of current detection methods. In particular, studies are being funded by WADA to identify a ‘molecular signature’ of rHumanEPO doping and preliminary results are promising. In the first systematic study to be conducted, the expression of hundreds of genes were found to be altered by rHumanEPO with numerous gene transcripts being differentially expressed after the first injection and further transcripts profoundly upregulated during and subsequently downregulated up to 4 weeks postadministration of the drug; with the same transcriptomic pattern observed in all participants. The identification of a blood ‘molecular signature’ of rHumanEPO administration is the strongest evidence to date that gene biomarkers have the potential to substantially improve the analytical performance of current antidoping methods such as the Athlete Biological Passport for rHumanEPO detection. Given the early promise of transcriptomics, research using an ‘omics’-based approach involving genomics, transcriptomics, proteomics and metabolomics should be intensified in order to achieve improved detection of rHumanEPO and other doping substances and methods difficult to detect such a recombinant human growth hormone and blood transfusions.
- Altitude
- Athletics
- Doping
- Elite performance
- Endurance
Statistics from Altmetric.com
Introduction
Erythropoietin
Erythropoietin is a glycoprotein hormone produced primarily in the kidneys that regulates red blood cell mass by stimulating the survival, proliferation and differentiation of erythrocytic progenitors.1 Anaemia is a common complication of chronic kidney disease caused principally by a deficiency in the synthesis of endogenous erythropoietin.2 Recombinant human erythropoietin (rHumanEPO) was first produced in the 1980s.3 ,4 In 1989, a 40-year-old patient with chronic kidney disease-associated anaemia who had undergone chronic haemodialysis for 7 years, an unsuccessful kidney transplantation and received 313 units of red blood cells, was the first patient to receive rHumanEPO treatment.5 Within 8 weeks of rHumanEPO therapy, his haematocrit increased from 15% to 38%, red blood cell transfusions were no longer required, he was able to play handball and resumed his physically demanding work.5 Since this success of almost biblical proportions, many clinical trials have demonstrated the efficacy and short-term safety of rHumanEPO for increasing and maintaining haemoglobin concentration, for reducing the need for red blood cell transfusions and improving the quality of life of patients with anaemia.6 These positive effects of rHumanEPO have to be weighed against the increased risk of venous thromboembolic events and potential unknown adverse long-term effects in patients.6 rHumanEPO is currently widely used to treat symptomatic anaemia caused by chronic kidney disease and anaemia with related symptoms in some patients with cancer receiving chemotherapy, as well as to increase the amount of blood that can be taken from individuals donating their own blood before surgery.
Brief history of drug detection in sport with particular reference to blood manipulation and erythropoietin
During the Olympic Games held at moderate altitude (2250 m at sea level) in Mexico City in 1968, altitude-induced blood adaptations such as an increase in haemoglobin concentration were considered primarily responsible for athletes living at altitude wining most of the endurance races and this phenomenon instigated a new research focus on altitude training in the field of sports physiology. As an example, the pioneering work of Ekblom et al7 demonstrated that elevation in haemoglobin concentration increased maximal oxygen uptake ( ) and improved performance. It is notable that the first accounts of blood manipulation in sport as well as the term ‘blood doping’ emerged soon thereafter in the media.8 There is evidence that blood transfusions were being used in the 1970s and 1980s by athletes such as cyclists, marathoners and cross-country skiers.9
) and improved performance. It is notable that the first accounts of blood manipulation in sport as well as the term ‘blood doping’ emerged soon thereafter in the media.8 There is evidence that blood transfusions were being used in the 1970s and 1980s by athletes such as cyclists, marathoners and cross-country skiers.9
Doping in sport seems to adapt rapidly to new available technology. The appearance of rHumanEPO, which was shown to successfully increase haemoglobin concentration in patients, radically transformed blood doping practices. Following the ‘transfusion era’, rHumanEPO doping took over in the 1990s, as was highlighted by numerous doping cases during the Tour de France cycle race and especially the Festina affair in 1998.10 In 2000, Birkeland et al11 demonstrated that rHumanEPO administration for 4 weeks in 20 male athletes increased haematocrit from 43% to 51%,  by 7% as well as time to exhaustion by 9%. The practice of red cell transfusions has made a strong comeback in recent years in response to the development of antidoping tests. Based on testimonies from athletes, it is now known that athletes are using rHumanEPO in combination with blood transfusions.10
by 7% as well as time to exhaustion by 9%. The practice of red cell transfusions has made a strong comeback in recent years in response to the development of antidoping tests. Based on testimonies from athletes, it is now known that athletes are using rHumanEPO in combination with blood transfusions.10
In 1990, erythropoietin was included on the list of prohibited substances by the International Olympic Committee because misuse by athletes was suspected, although no approved test existed.12 There were rumours during this time that athletes used large doses of rHumanEPO to induce very high-blood parameter values. For example, Bjarne Riis, the 1996 Tour de France winner (who admitted doping in 2007), acquired the nickname of ‘Mr 60%’ which referred to his haematocrit level.13 Because rHumanEPO was directly undetectable at the time and as well as to protect the health of athletes, the 50% haematocrit rule was introduced in 1997 by the International Cycling Union.14 Briefly, any racing cyclist who had a haematocrit above 50% (50% for male athletes and 47% for female athletes) was declared ineligible and was excluded from the race. Despite this rule, the Festina scandal at the Tour de France in 1998 provided the proof of organised and widespread doping in professional cycling and highlighted the need for the creation of an independent international agency, which would set unified standards for antidoping work and coordinate the efforts of sports organisations and public authorities. The WADA was consequently established in 1999.
Although the detection of rHumanEPO doping is difficult because rHumanEPO is structurally very similar to endogenous erythropoietin, a direct urinary test based on the presence of different isoforms in isoelectric profiles was developed and published by Lasne et al.15 This test became the first approved method for the direct detection of rHumanEPO doping. While the effects of rHumanEPO administration can last for weeks, the test has a rather limited detection window, that is, a few hours up to a few days depending on the dose used, the route of administration and the individual metabolism.16 ,17 At the same time as the development of the direct test, potential indirect detection of rHumanEPO using blood markers of altered erythropoiesis was being investigated.8 Notably, following the initial work of Parisotto et al18 (ie, the so-called first generation blood tests), Gore et al19 developed the second generation of statistical models using indirect markers of altered erythropoiesis in order to detect rHumanEPO doping during (ON-model) and after repeated recent (OFF-model) administration. However, despite the promising laboratory-based results, both direct and indirect detection methods are not immune from false negatives17 ,20 ,21 or indeed false positives.18 ,22 For example, it is known that terrestrial or simulated altitude exposure can influence the indirect indices of rHumanEPO doping.23–25 Despite considerable efficacy, there was clearly a need for these original detection methods to be improved.
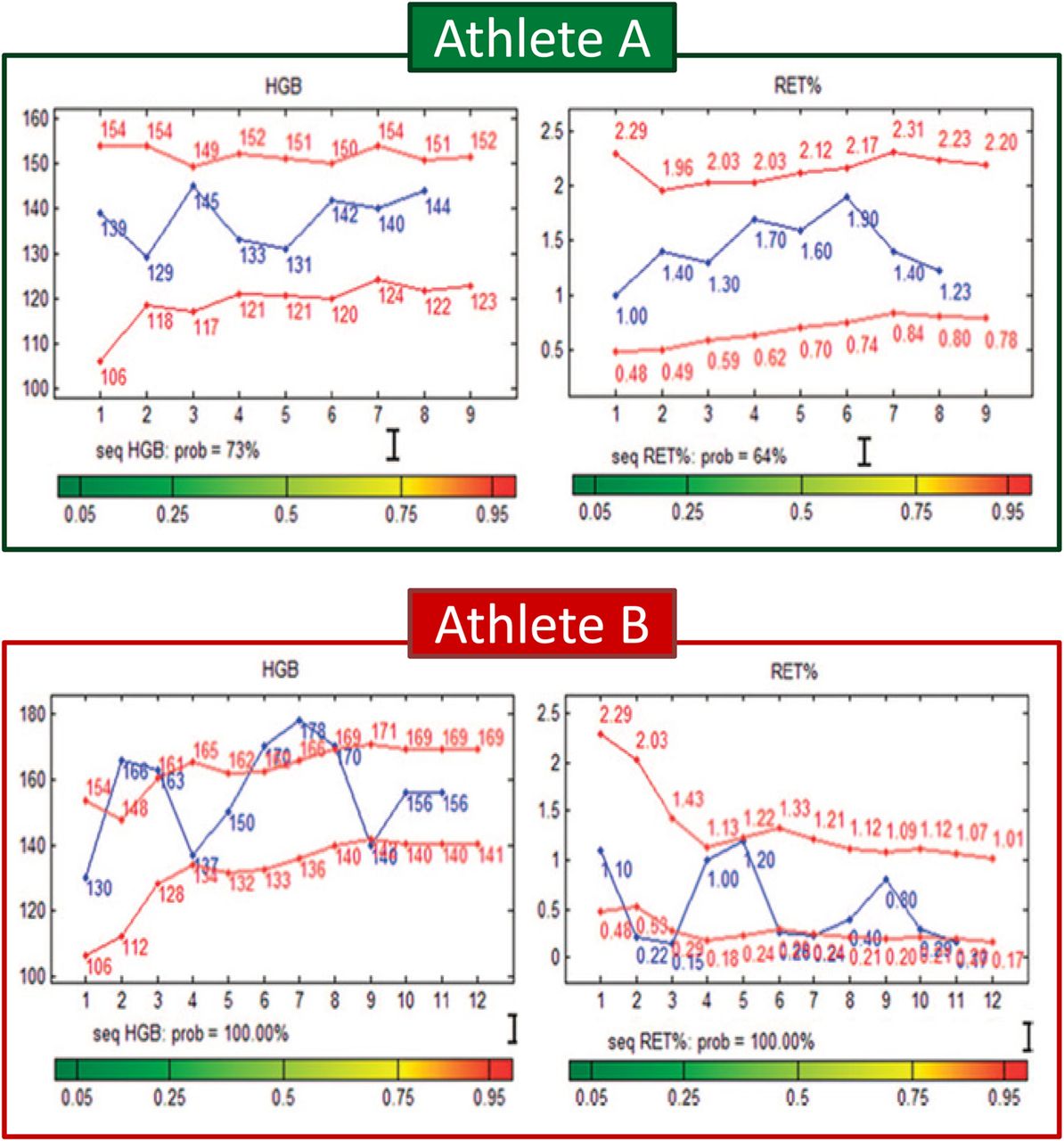
The Athlete Biological Passport (ABP) was recently introduced as a new tool to indirectly detect erythropoiesis-stimulating agents such as rHumanEPO, which can lead to a doping sanction and at the same time as intelligently targeting athletes for additional testing.26 The ABP approach relies on identifying intraindividual abnormal variability over time in selected haematological parameters (figure 1).14 ,27 The blood doping behaviour of athletes has changed since the introduction of the ABP.28 Extreme reticulocyte values (ie, <0.4% or >2%) have dramatically decreased and as such, the implementation of the ABP has undeniably been a step forward in the antidoping field.28 Although the ABP approach can identify abnormal enhanced erythropoiesis regardless of the method used, such as autologous blood transfusion, as shown by Pottgiesser et al29 in a field-like longitudinal blinded setting, the sensitivity of the ABP has been questioned.30 In order to minimise the risk of being caught via the ABP, it is well known that some athletes are now using ‘microdoses’ of rHumanEPO which allegedly range from 10 to 40 IU/kg body mass.16 ,20 ,31 Microdoses of rHumanEPO aim (1) to increase haemoglobin mass (Hbmass) while avoiding large fluctuations in the ABP blood markers as well as minimising the detection window for conventional direct methods and/or (2) to ‘normalise’ the ABP blood markers after blood manipulations such as autologous blood transfusion.20 ,31 In addition, variation in ABP haematological parameters due to factors such as training or hypoxia exposure can influence the interpretation of the ABP results.32 Adjustments by the athletes to antidoping methods impose a constant need for scientists and antidoping authorities to develop and implement new detection methods based on innovative concepts and cutting-edge technologies.

Examples of the Athlete Biological Passport (ABP) including haemoglobin concentration (HGB) and reticulocyte percentage (RET%). Blue lines: actual tests results. Red lines: individual ‘normal’ limits calculated by the ABP software. Athlete A presented normal variations. Conversely, athlete B presented abnormal variations, and was consequently suspected of doping. Adapted from Sottas et al.27
‘Omics’-based approaches to detect rHumanEPO doping
The initial ‘omics’ studies
A genome is defined as the entire collection of genetic information encoded by a particular organism.33 By semantic association, the words transcriptome, proteome and metabolome refer to the entirety of all transcripts, proteins and metabolites expressed by a genome at a specific time. The science related to these terms was named accordingly genomics, transcriptomics, proteomics and metabolomics and collectively encompass the so-called omics cascade34 (figure 2). Using microarray technology, a characteristic gene expression signature of a particular stimulus can be revealed by assessing the transcription state of the cells of the experimental samples.35 Therefore, if the gene expression profile following rHumanEPO administration yields a specific gene expression signature, this could lead to the development of new methods with potentially improved discriminatory power relative to current detection protocols. Some initial support for this hypothesis emerged in a WADA funded study by Varlet-Marie et al.36 Using the serial analysis of gene expression (SAGE) method, Varlet-Marie et al identified 95 genes whose differential expression was subsequently tested by quantitative real-time PCR in two athletes. These athletes were treated first with high doses of rHumanEPO and then with microdoses. Thirty three marker genes for rHumanEPO administration were identified during the high-dose regimen and five remained differentially expressed during the microdose regimen. A transcriptomic-based approach has also been applied with some success to other doping methods and substances used by athletes. For example, characteristic gene expression profiles have been shown in six patients after autologous blood transfusion.37 Collectively, these pilot studies, involving only a limited number of patients, provide some confirmatory data to support the hypothesis that gene expression profiles may provide a sensitive method for the detection of rHumanEPO and blood doping.

The ‘omics’ cascade. The cascade depicts the flow of information from gene to metabolism.
To build on the initial promise and develop new methods with improved discriminatory power relative to current detection protocols using ‘omics’ technologies, a series of studies have been funded by WADA as follows:
-
A gene-microarray-based approach to the detection of recombinant human erythropoietin doping in endurance athletes.
-
Application of a minimally invasive method for RNA sampling and the addition of miRNA to the detection of recombinant human rHumanEPO use by athletes.
-
Metabolomic profiling of recombinant erythropoietin rHumanEPO in Caucasian and east-African endurance-trained athletes.
-
A systems biology biomarker-based approach to the detection of microdose recombinant human erythropoietin doping.
-
A new dimension to the detection of recombinant human erythropoietin doping using blood gene expression profiles: development of direct antidoping applications.
-
A systems biology biomarkers approach to the differentiation of recombinant human erythropoietin doping from confounding factors.
The overall aim of these studies was to identify a molecular signature of rHumanEPO doping and hence provide a basis for the development of novel and robust testing models to detect rHumanEPO doping. Using both phenotypic and molecular methods, the first study was carried out at sea level in Glasgow (Scotland (SCO), UK) with Caucasian endurance-trained men38 and at moderate altitude (∼2150 m) in Eldoret (Kenya (KEN)) with KEN endurance runners39 who abstained from official sporting competition for the entire duration of the study. The specific aims of this first study were:
-
To determine and compare the effects of rHumanEPO administration on blood parameters and exercise performance in endurance-trained men living and training at or near sea level with another cohort living and training at moderate altitude (∼2150 m) in KEN.
-
To establish adequate molecular methods and molecular workflow to enable valid and optimal assessment of blood gene expression.
-
To assess the effects of rHumanEPO on blood gene expression profiles in endurance-trained volunteers living and training at or near sea level.
-
To replicate and compare the effects of rHumanEPO administration on blood gene expression profiles in another cohort living and training at moderate altitude (∼2150 m) in KEN.
-
To validate the gene-microarray-based findings using another specific and sensitive quantitative gene expression technology.
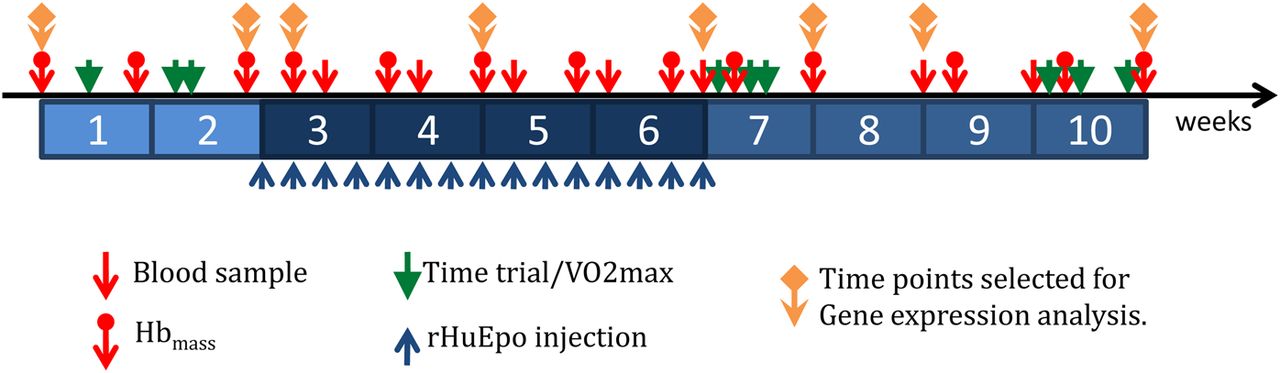
To achieve these ambitious aims, 39 endurance-trained men, 19 based at sea level (Glasgow, SCO) and 20 based at moderate altitude (Eldoret, KEN, 2100–2800 m at sea level), received rHumanEPO injections for 4 weeks. Blood was obtained 2 weeks before, during and 4 weeks after administration (figure 3). Blood, urine and saliva samples at 20 time points per volunteer (780 blood samples ie, 20 time points×39 participants; figure 3) were collected and specifically processed and stored at −80°C for current and future exploitation. Time points at key stages were selected for gene expression analysis (figure 3). Phenotypic results from this first study are beginning to emerge.38 ,39 Briefly, relative to baseline, running performance was significantly improved following 4 weeks of rHumanEPO administration in sea level Caucasian-trained men and remained significantly elevated 4 weeks after administration by approximately 6% and 3%, respectively; these performance effects coincided with significantly rHumanEPO-induced elevated  and Hbmass.38 rHumanEPO administration improved
and Hbmass.38 rHumanEPO administration improved  and time trial performance to a similar extent in KEN and SCO runners despite a higher haematocrit and haemoglobin concentration in KEN compared with SCO prior to rHumanEPO and similar at the end of administration.39 With regard to the transcriptomic data, preliminary results show that relative to baseline, hundreds of genes were differentially expressed during rHumanEPO administration.40 Notably, a few of these genes were already upregulated after only one single rHumanEPO injection, while further genes were differentially expressed during rHumanEPO administration and remained differentially expressed up to 4 weeks after administration compared with baseline. In summary, a signature pattern of several genes upregulated during rHumanEPO and subsequently downregulated up to 4 weeks after administration in the SCO cohort.40 More recent transcriptomic data to emerge from this study show replication of the SCO microarray data in the KEN cohort.41 An example of the characteristic gene expression pattern in the sea level Caucasian and Kenyan altitude-based cohorts is illustrated in figure 4. A subset of target genes was further validated using another quantitative gene expression technology.41 From the preliminary data, it is clear that significant progress has been made towards successfully fulfilling all five objectives. This research has successfully identified, replicated and validated the blood ‘molecular signature’ of rHumanEPO administration and as such provides the strongest evidence to date that gene biomarkers have the potential to improve the performance of current antidoping methods such as the ABP for rHumanEPO detection.
and time trial performance to a similar extent in KEN and SCO runners despite a higher haematocrit and haemoglobin concentration in KEN compared with SCO prior to rHumanEPO and similar at the end of administration.39 With regard to the transcriptomic data, preliminary results show that relative to baseline, hundreds of genes were differentially expressed during rHumanEPO administration.40 Notably, a few of these genes were already upregulated after only one single rHumanEPO injection, while further genes were differentially expressed during rHumanEPO administration and remained differentially expressed up to 4 weeks after administration compared with baseline. In summary, a signature pattern of several genes upregulated during rHumanEPO and subsequently downregulated up to 4 weeks after administration in the SCO cohort.40 More recent transcriptomic data to emerge from this study show replication of the SCO microarray data in the KEN cohort.41 An example of the characteristic gene expression pattern in the sea level Caucasian and Kenyan altitude-based cohorts is illustrated in figure 4. A subset of target genes was further validated using another quantitative gene expression technology.41 From the preliminary data, it is clear that significant progress has been made towards successfully fulfilling all five objectives. This research has successfully identified, replicated and validated the blood ‘molecular signature’ of rHumanEPO administration and as such provides the strongest evidence to date that gene biomarkers have the potential to improve the performance of current antidoping methods such as the ABP for rHumanEPO detection.
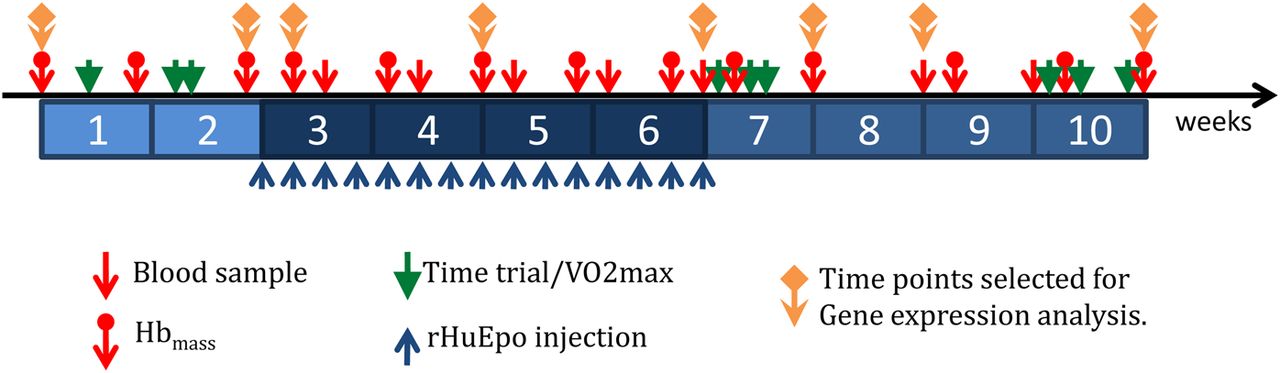
Experimental design for the WADA project entitled ‘A gene-microarray-based approach to the detection of recombinant human erythropoietin doping in endurance athletes’. Hbmass, haemoglobin mass; rHuEpo, recombinant human erythropoietin; VO2max, maximal oxygen uptake.

A typical gene expression pattern for illustrative purposes. EPO, erythropoietin; KEN, Kenya; SCO, Scotland
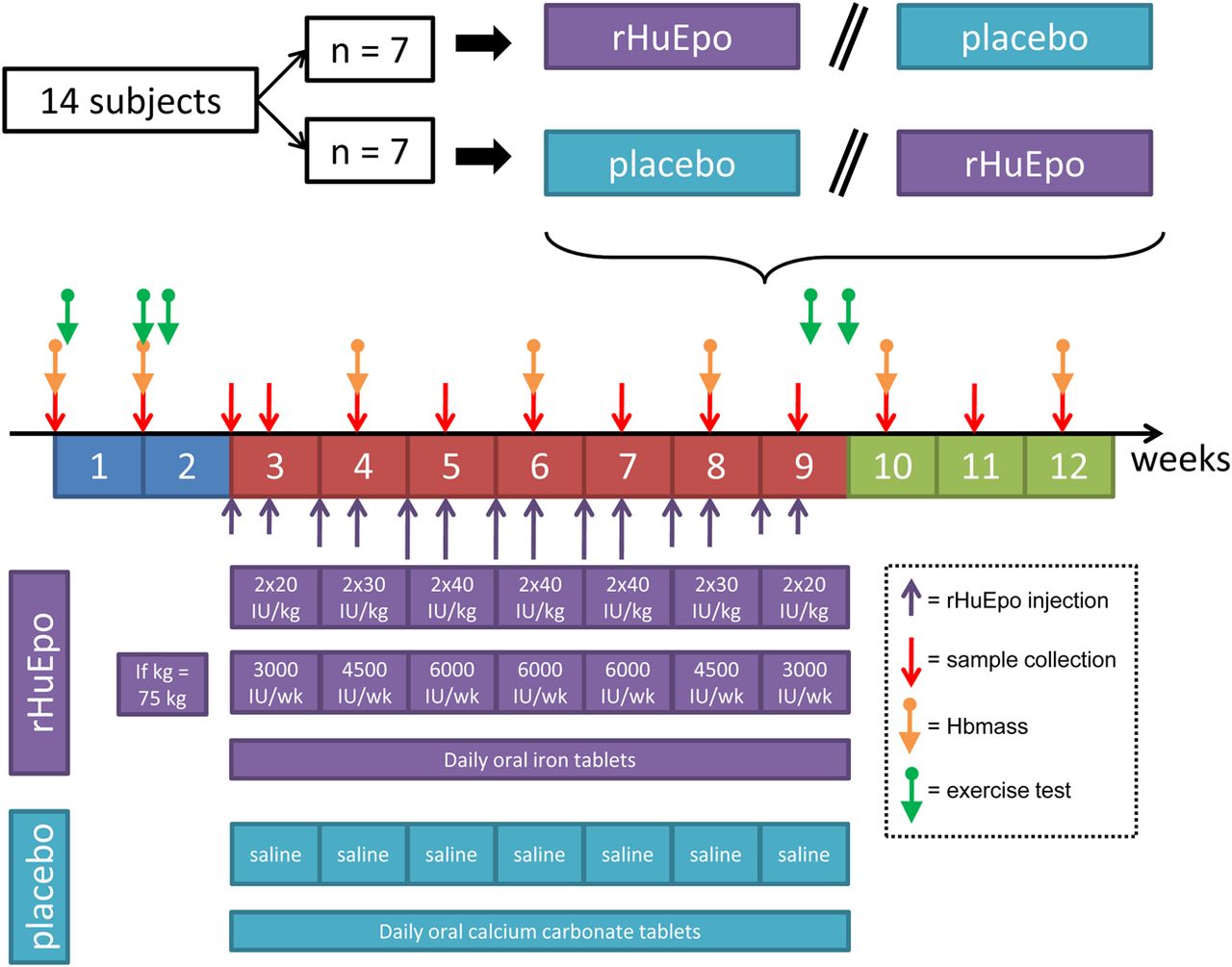
The results from the first study are promising, however, it is well recognised that some athletes are now microdosing with rHumanEPO to minimise the risk of being caught via currently applied detection methods.16 ,20 Interestingly, five genes remained differentially expressed even when high and microdoses were taken into account together in the study by Varlet-Marie et al.36 While these results were generated using SAGE and from only a few participants, they provide some confirmatory data to support the idea that gene expression profiles may provide a more sensitive method to detect microdoses of rHumanEPO, especially when the whole genome is interrogated. Therefore, a second study was funded by WADA and designed to use the genes significantly altered using a fairly high regimen of rHumanEPO injections discovered in the first study as primary detection candidates of microdosing with rHumanEPO. For this currently ongoing study, 14 healthy endurance-trained individuals not involved in competition were required to participate in a randomised, double-blind, placebo-controlled crossover microdose rHumanEPO regimen necessitating the collection of 364 blood samples (ie, 13 samples×14×2; figure 5). The aim of the rHumanEPO microdose regimen was to increase Hbmass while deliberately avoiding large fluctuations in reticulocyte production. Briefly, each participant received rHumanEPO injections (NeoRecormon, Roche, Welwyn Garden City, UK) subcutaneously twice per week for 7 weeks (figure 5). Blood samples were collected in triplicate at baseline and then once a week during rHumanEPO administration and for 3 weeks after rHumanEPO administration (ie, 13 blood samples per participant; figure 5). Standardised collection and storage procedures were applied as per the previous study.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Experimental design for the WADA project entitled ‘A systems biology biomarker-based approach to the detection of microdose recombinant human erythropoietin doping’. Hbmass, haemoglobin mass; rHuEpo, recombinant human erythropoietin.
Current confounder studies
Given the ongoing studies and promising preliminary data, it is of paramount importance to evaluate the effects of major confounding factors on the discovered ‘molecular signature’ of rHumanEPO doping, such as the effects of altitude. It is known that natural or simulated altitude training is used by athletes for performance enhancement,42 although the optimal regimen and magnitude of ergogenic effect of natural altitude training is unclear, while there is a small/no ergogenic effect of simulated altitude.43 Only the live high –train low method developed by Levin and Stray-Gundersen44 using natural altitude would appear to be effective.43 In addition, it remains uncertain to what extent these ergogenic effects are due to augmented erythropoiesis.43 Regardless of these conflicting results, altitude exposure may potentially influence haematological indices of rHumanEPO doping.23 ,24 As a result, some athletes claim altitude exposure as a source of variation of their individual blood variables to mask blood doping practices such as the administration of rHumanEPO. There is therefore a need to develop specific and robust testing models which can differentiate altitude training from blood doping combined (or not) with altitude exposure and preliminary unpublished data using transcriptomics are promising (WADA unpublished data). Therefore, the aim of a third study recently funded by WADA is to compare blood gene expression profiles altered by rHumanEPO (fairly high-dose and microdose regimens, data from past and ongoing WADA-funded projects) with altitude exposure in order to provide a set of candidate genes that can be used to differentiate rHumanEPO from altitude training. For this study, at least 20 endurance athletes will spend at least 4–6 weeks at altitude above 2000 m on more than one occasion and serial blood samples collected prior, during and post altitude exposure. The precise timing of blood sample collection will be determined from pilot altitude studies currently underway. Once again, standardised blood collection and storage procedures will be applied as per previous studies and in doing so add further important samples to the ‘omics’ biobank being created for current and future doping test development.
In addition to altitude, another major potential confounding factor that could impact on the ‘molecular signature’ of rHumanEPO doping is the effect(s) of prior exercise. Samples are often collected at sporting or training venues after intense exercise.45 It has previously been demonstrated that exercise significantly affects gene expression profiles of peripheral blood mononuclear cells and white cells count.46 ,47 It is essential, therefore, to define the effects of exercise on blood gene expression profiles and particularly on the ‘molecular signature’ of rHumanEPO doping. In order to provide a set of robust candidate genes that can be used for the detection of rHumanEPO doping, blood gene expression assessment will be performed during and post intense exercise after microdose rHumanEPO or placebo in the double-blinded trial (ie, WADA study 2, figure 5). In particular, a modified Wingate test consisting of 10 sprints of 10 s at baseline (following a familiarisation) and during the week after microdose rHumanEPO (figure 5) in order to provide a set of robust candidate genes that can be used for the detection of rHumanEPO doping but also examine the effects of rHumanEPO on progressive anaerobic fatigue and repeated sprint ability. The ability to reproduce maximal performance in subsequent short-duration sprints separated by brief recovery periods is a key fitness requirement in most team and racket sports.48 Of note, the International Tennis Federation recently adopted the ABP programme, while there is anecdotal evidence in football that doping may be higher than perceived by sport authorities.49 ,50 In addition, these results will be used to provide the basis for the development of recommendations on the collection of samples in order to ensure the ‘validity’ of the blood gene expression results. It is hoped that this approach will help reduce the pressure on the athletes who need to wait 2 h after training or competition before a blood sample is taken for the ABP.
Necessary steps and potential future applications of an ‘omics’-based approach to detect rHumanEPO doping
The ABP approach relies on identifying intraindividual abnormal variability over time of selected haematological parameters.14 More precisely, the ABP approach uses Bayesian networks for the evaluation of the likelihood of doping based on several variables and/or factors which can then be used as evidence for disciplinary sanction. Bayesian networks are interpretable and flexible models for representing the probabilities of causal relationships between multiple interacting variables.14 ,51 In the antidoping field, the Adaptive Model based on the Bayesian approach directly estimates the probability of blood doping based on previous individual test history and heterogeneous factors known to influence blood parameters. This feature allows the model to remove the variance due to interindividual differences and heterogeneous factors such as gender, ethnicity, altitude exposure, age and sport discipline.51 Based on the promising transcriptomic findings to date, blood gene biomarkers can potentially be integrated into the ABP in order to further improve its analytical performance. The idea being to build on the current ABP approach by combining currently used haematological parameters with new molecular signature data in the first instance and in doing so attempt to create a unique ABP model with improved specificity and sensitivity, not amenable to manipulation.
In order to evaluate the proof of principle concept outlined here, interindividual and intraindividual variability as well as factors which can affect blood gene expression profiles must be precisely determined. It has been previously demonstrated that external factors can influence blood profiles. For instance, strenuous endurance exercise can induce changes in some blood parameters not only by the well-described increase in plasma volume known as the athlete's pseudoanaemia52 but also by exercise-induced haemolysis due to metabolic modifications, oxidative stress and repeated foot impact (foot strike).53–55 The effects of strenuous long distance exercise such as cycling stage racing and distance running on blood gene expression profiles should consequently be investigated in elite endurance athletes. As most blood parameter values differ substantially between men and women, the currently used reference ranges in the ABP were determined separately for each sex.51 ,56 A similar differentiation approach could also be applied to blood gene expression profiles in order to evaluate gender differences and similarities. Injury-related immobility also has the potential to affect blood gene profiles, as it has been reported that immobility for a period of 4 weeks reduces Hbmass by ∼19% in a competitive international female athlete.57 Research to date has demonstrated that ethnicity and residence at altitude only minimally influence the blood gene expression signature of rHumanEPO doping. However, more research is required to measure and generate normal reference ranges and the impact of other ‘confounders’ on blood gene expression profiles to enhance further the discriminant power of the ABP. If these key follow-up studies are successful, and an ‘omics’-based solution is adopted in the future, one option would be to adopt on mass (eg, full squad/team), announced testing to obtain baseline data that could also be used as a comparison throughout the season and for subsequent seasons, with target testing set up on any possible anomalies; such testing could even be initiated at youth/academy level to build up profiles and reference ranges to be monitored over time. However, in this case, possible age-related changes in the reference range of the selected molecular signatures of doping would need to be considered. In addition to determining the biological variability of ‘omics’ measures, markers and confounders, successful implementation of an ‘omics’ antidoping solution will require the analytical variability of the finally chosen ‘omics’ technology to be well established given the importance of analytical and preanalytical factors in any antidoping rule violation procedure. The true potential of ‘omics’ technology and antidoping application (eg, potential to reduce the number of traditional antidoping tests) will require this essential process to be undertaken.
Conclusions
The current research on the molecular signature of rHumanEPO doping has, so far, provided some evidence that ‘omics’ technologies have the potential to significantly strengthen the ABP approach and contribute to other traditional antidoping tests. This approach can be applied for detection of other doping substances and methods difficult to detect such a recombinant human growth hormone and blood transfusions. There is also the interesting possibility that this approach could help reducing the pressure on the athletes’ antidoping obligations such as the ‘athletes whereabouts’. In order to confirm that an integrative ‘omics’ approach is a solution to improve rHumanEPO detection, it is of paramount importance to precisely determine the normal reference values of the blood molecular signature of rHumanEPO doping using different dose regimens as well as to carefully assess the potential effects of external factors on blood gene expression profiles, such as prior training, altitude including different hypoxic ‘dose’ and protocols, sport discipline, level of competition, gender, ethnicity and age. These investigations are necessary before potentially including the promising blood gene biomarkers in the ABP and/or the development of a stand alone test to reveal doping or identify suspicious samples for targeting purposes.
Acknowledgments
The authors wish to thank the members of the steering committee of their WADA-funded research for their valuable comments and the numerous students who assisted data collection in Scotland and Kenya. The cooperation of all the participants is also greatly appreciated.
References
Footnotes
-
Funding The series of studies reviewed here were all supported by research grants from the WADA.
-
Competing interests None.
-
Provenance and peer review Commissioned; externally peer reviewed.