Article Text

Abstract
A model is proposed in which the development of physical exhaustion is a relative rather than an absolute event and the sensation of fatigue is the sensory representation of the underlying neural integrative processes. Furthermore, activity is controlled as part of a pacing strategy involving active neural calculations in a “governor” region of the brain, which integrates internal sensory signals and information from the environment to produce a homoeostatically acceptable exercise intensity. The end point of the exercise bout is the controlling variable. This is an example of a complex, non-linear, dynamic system in which physiological systems interact to regulate activity before, during, and after the exercise bout.
- EMG, electromyographic
- IEMG, integrated electromyographic
- MVC, maximal voluntary contraction
- teleoanticipation
- fatigue
- brain
- neural recruitment
- pacing strategies
Statistics from Altmetric.com
Fatigue is usually defined as a decrease in force production,1,2 or an inability to regenerate the original force3 in the presence of an increased perception of effort.4 The physiological causes of fatigue have been classified as either “peripheral” or “central” in origin. Peripheral skeletal muscle fatigue is usually defined as a decrease in the capacity of the skeletal muscle to generate force because of action potential failure, excitation-contraction coupling failure, or impairment of cross bridge cycling, in the presence of unchanged or increasing neural drive.5,6 In contrast, central fatigue has been defined as a reduction in neural drive to the muscle resulting in a decline in force production or tension development that is independent of changes in skeletal muscle contractility.4
Metabolite changes believed to cause peripheral fatigue during maximal short term voluntary contractions include increased lactic acid concentrations, pH decreases and associated proton accumulation, depletion of ATP and creatine phosphate, accumulation of ADP, IMP, and inorganic phosphate,7 changes in the skeletal muscle Na+/K+ ATPase pump, and sarcolemmal, t-tubule, and sarcoplasmic reticulum Ca2+ mediated functional changes.8–14
Similarly, it is suggested that during progressive maximal exercise to exhaustion, the inability to increase oxygen consumption with resultant skeletal muscle anaerobiosis,15,16 or excessive heat accumulation17,18 causes fatigue.19–22 During submaximal endurance exercise, the depletion of muscle or liver glycogen stores is believed to cause fatigue.23–28
According to the peripheral model of fatigue, these metabolic changes lead to system failure of either the peripheral muscles or the heart, causing the termination of exercise even though there is increased central neural drive from the brain to the peripheral muscles.5,6 The supposed goal of the increasing neural drive is to maximise skeletal muscle recruitment allowing exercise to continue. However, the clearly deleterious effect would be to exacerbate the metabolic crisis that, according to this model, causes the termination of exercise. Once there is maximal muscle recruitment, the terminal metabolic crisis ultimately causes skeletal muscle contractile failure according to the catastrophe model of Edwards.29 Thereafter there is an absolute requirement for a period of rest while the metabolic crisis is reversed and the concentrations of inhibitory metabolites return to normal, allowing a full functional recovery.
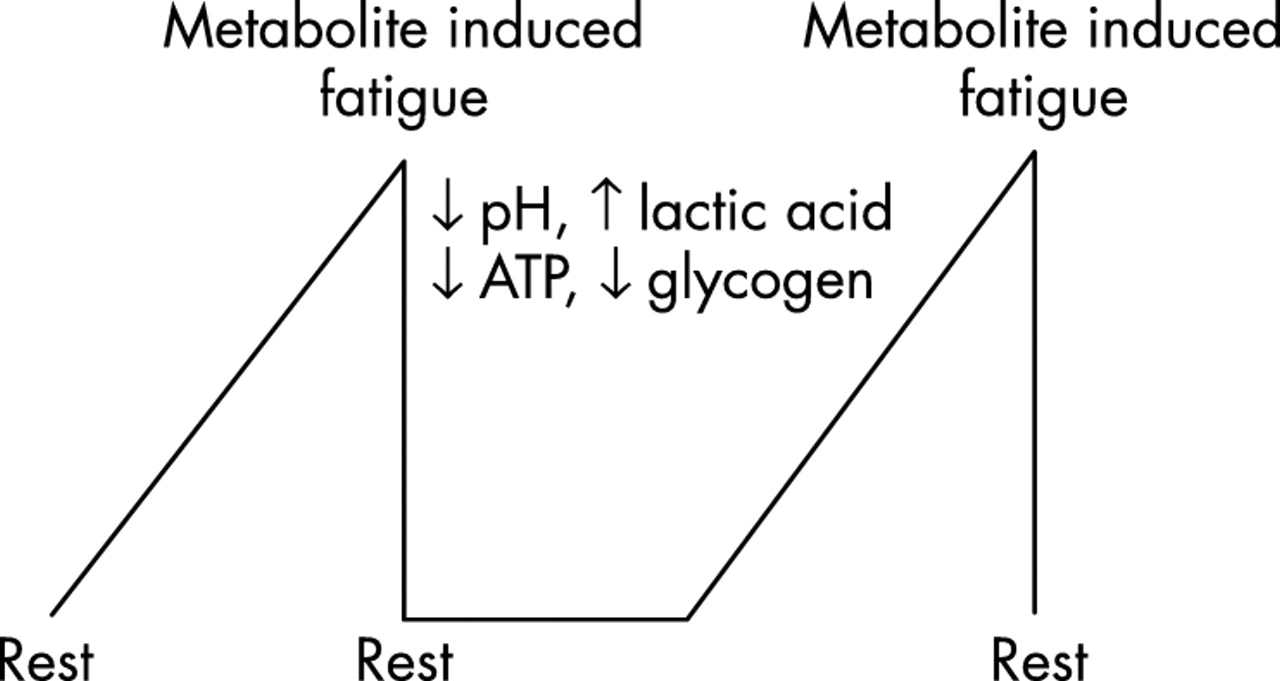
The peripheral fatigue model therefore predicts that exercise always terminates at an absolute, temporarily irreversible end point. Such an absolute physiological end point is an example of a linear dynamic system30 in which changes occur in direct response to the input variable. The maximal and minimal capacities of the system are related to the minimal and maximal capacity of the input variable. Therefore, according to this model, a subject who begins fatiguing exercise will continue until an absolute end point of exhaustion is reached (fig 1). The timing of this end point will depend on the maximal capacity of the limiting physiological and metabolic processes. Only after a suitable period of complete inactivity during which the processes causing fatigue are reversed, does further activity again become possible31,32 (fig 1). In this peripheral model therefore fatigue is a negative and unavoidable consequence of physical activity.33 This model has proved the most popular explanation for fatigue for the past several decades.34

Peripheral fatigue as usually conceived is an example of a linear dynamic system in which the progressive accumulation of metabolites in the exercising muscles causes an absolute fatigue, after which a period of rest is required before exercise can again be undertaken. This model also predicts that, at the point of absolute fatigue, the brain has recruited all the motor neurones in the active skeletal muscles.
However, when used to explain the fatigue of human exercise, certain predictions of the linear dynamic model are not demonstrable. Firstly, absolute skeletal muscle substrate depletion or metabolite accumulation has only been shown in muscle tissue studied in vitro or in intact skeletal muscle stimulated to contract artificially.32,35 However, none of these absolute metabolic changes has been shown directly to cause “fatigue” during either static or dynamic exercise in vivo in the presence of an intact central nervous system.8,34,36–40
Secondly, according to this model, the sensations of fatigue must convey a conscious perception of the physiological and metabolic changes in the peripheral organs, most especially the active skeletal muscles. Thus in this model, metabolic and other changes in the peripheral organs progressively impair skeletal muscle contractile function, and the sensation of fatigue is a sensory copy of those peripheral changes (fig 2).41
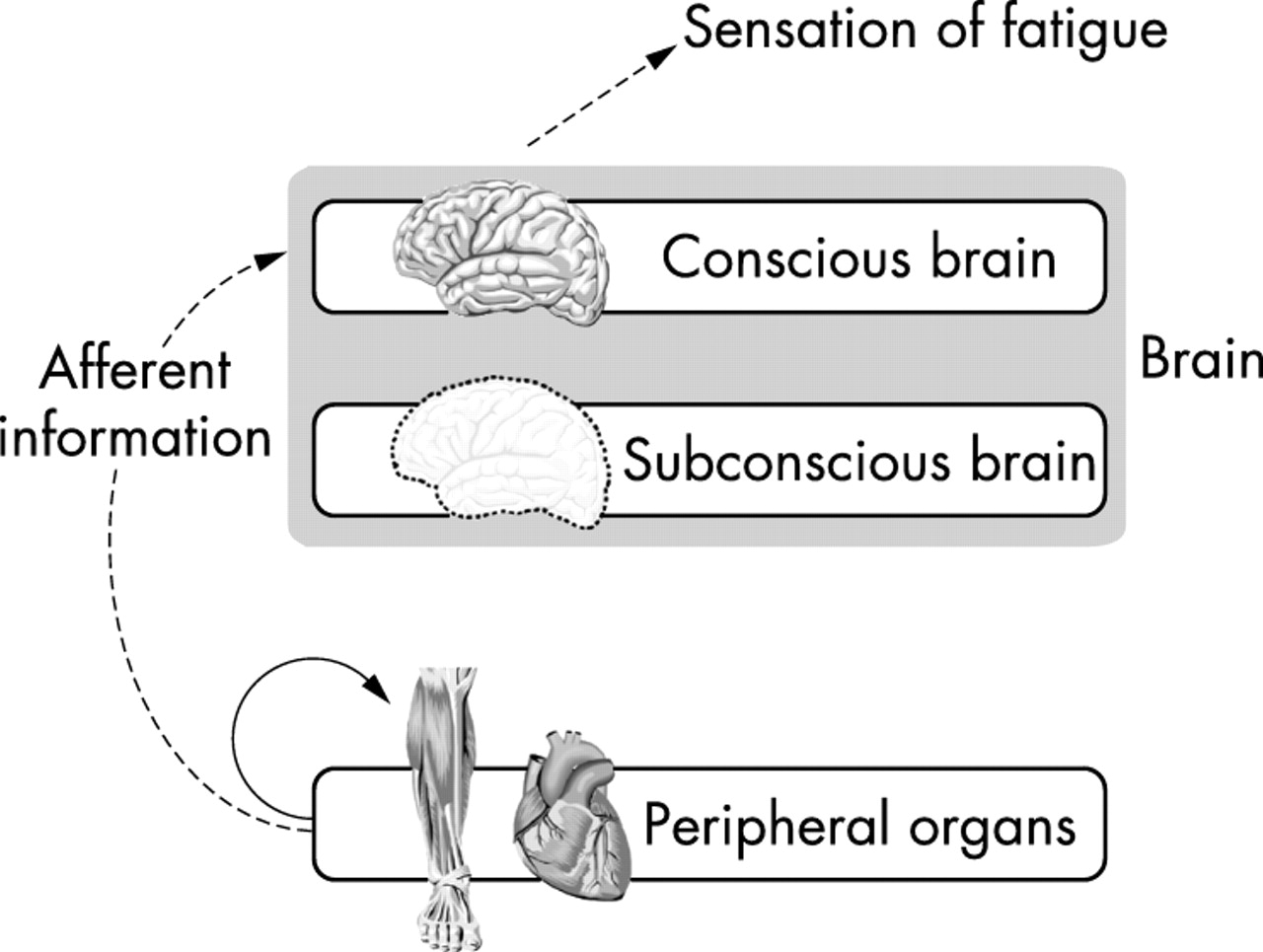
Accumulation of fatiguing metabolites in the active muscles impairs skeletal muscle contractile function according to the model of peripheral fatigue. The same metabolic changes would also elicit the sensations of fatigue. However, these sensations can have no value because fatigue is peripherally based so that the muscles must stop working regardless of any input from the brain.
However, no study has yet clearly established a direct relation between any single physiological variable and the perception of effort or fatigue.41–49 In contrast, several studies have shown the opposite. For example, patients with chronic fatigue syndrome experience fatigue even at rest and participate in any physical activity with reluctance.50,51 Yet the exercise capacity of patients with this condition is not impaired52–54 so that the physiological bases for their symptoms remain unknown. However, the “mismatch” between the severity of their symptoms despite an unimpaired exercise capacity indicates that their fatigue must be associated with alterations in neural drive55 and originates in those brain areas responsible for the conscious perception of effort and fatigue, rather than for motor control.4,52 These findings confirm that the conscious sensation of fatigue is not always directly related to the exercise intensity.56
Patients who are pyrexial also experience symptoms of fatigue at rest.57 Cytokines released from the immune cells during the pyrexial illness may activate the conscious perception of fatigue, the function of which would be to speed recovery58 by avoiding exercise. Therefore, in this condition, the increased sense of fatigue even without physical activity has survival value because it acts to prevent unnecessary physical activity.
Accordingly as the linear, peripheral model of fatigue is unable to explain certain fundamental phenomena, some other mechanism must be operative to prevent the development of absolute physical exhaustion while at the same time causing the conscious perception of the sensation of fatigue.
MODELS OF CENTRAL FATIGUE
The central fatigue model posits that the reduction in power output during prolonged exercise leading to the cessation of exercise is not caused by limiting physiological processes in any of the peripheral organs including the active skeletal muscles. Rather, the fatigue related reductions in skeletal muscle power output are caused by altered efferent command from the brain.59–63
One hypothesis is that changes in the concentration of certain brain neurotransmitters cause fatigue. For example, serotonin (5-hydroxytryptamine) concentrations increase during exercise as fatigue develops.64–70 Similarly, dopamine71–73 and acetylcholine74 concentrations decline, while concentrations of cytokines58 and ammonia75 increase. Inoue et al76 propose that the concentrations of chemicals such as actively transforming growth factor β increase in the cerebrospinal fluid during exercise and may also cause fatigue.
It is not clear if changes in the concentrations of these neurotransmitters are caused by the increased production in the peripheral tissues of their precursors, which then cross the blood-brain barrier,64,75 or whether they result from increased neural activity.
However, this hypothetical explanation also represents a model of “peripheral” fatigue, in which increases or decreases in neurotransmitter concentrations cause fatigue according to a linear model (fig 1). In this linear model, the increased neural concentration of neurotransmitters would be the “toxic” byproduct of neural activity as would be the increased concentration of other metabolites in the cerebrospinal fluid. Their increasing concentrations would subsequently impair the brain’s capacity to recruit an appropriate number of motor units. Thus any conscious desire to increase the power output by increasing motor unit recruitment would be blocked by these metabolic changes in the brain. This is analogous to the postulated action of lactate on skeletal muscle according to Hill’s cardiovascular/anaerobic model.77
The alternative model of central fatigue proposes that fatigue is not caused by these inhibitory metabolic processes in the central nervous system, but is part of an active neural process, the function of which is to prevent the development of absolute fatigue and organ damage, as proposed in the companion paper.77
Most researchers investigating central fatigue measure electromyographic (EMG) changes in the active muscles during submaximal or maximal contractions78–80 on the assumption that these changes reflect alterations in central (brain) neural drive. If fatigue is due to peripheral changes in the exercising skeletal muscles, then their EMG activity should increase progressively to compensate for the impaired contractile function caused by the defined metabolic changes (fig 3).4,5,81,82 The increased neural drive is therefore secondary to the progressively impaired contractile function of the exercising muscles.

Different patterns of integrated electromyographic (IEMG) changes during exercise indicating the development of peripheral and central fatigue. If fatigue is determined only by peripheral metabolic changes, then the work of the active skeletal muscles will fall progressively, whereas IEMG activity will either remain unchanged or increase. This indicates that the force output from individual muscle fibres is falling. In contrast, if fatigue is due to centrally mediated changes in the number of skeletal muscle motor units that are recruited, then both the work produced by the active skeletal muscles and the IEMG activity will fall in parallel, indicating that the recruitment of fewer motor neurones has caused the work output of the muscles to fall.
In contrast, the central fatigue model predicts that EMG activity falls during prolonged exercise as a result of a reduced central neural drive. Thus, in this model, the reduced skeletal muscle contractile function is secondary to a reduction in central neural drive (fig 3). Other factors that may influence the interpretation of the EMG during exercise83 fall beyond the scope of this review.
A number of studies have established that the decreased force production during muscular contractions may result from a reduced central neural drive and recruitment of skeletal muscle motor units secondary to inhibitory influences in response to afferent sensory information coming from the exercising muscles, the heart, or other peripheral organs.3,61,84–91 Other studies propose that this reduced motor unit recruitment may be caused by inhibitory reflexes stimulated by sensory afferents from group III or IV metaboreceptors in muscle acting at the level of the spinal cord.92–94
However, other studies have shown that altered central drive does not originate from the motor cortex or more “downstream” efferent neural structures.61,95–98 Rather, changes in motor command were found to be regulated by brain cortical processes “upstream” from the motor cortex, perhaps in areas such as the prefrontal and cingulate cortex. Alterations in neural drive during exercise could therefore result from commands generated in these higher cortical regions in response to somatosensory input from afferent fibres from peripheral organs such as the heart or skeletal muscles.
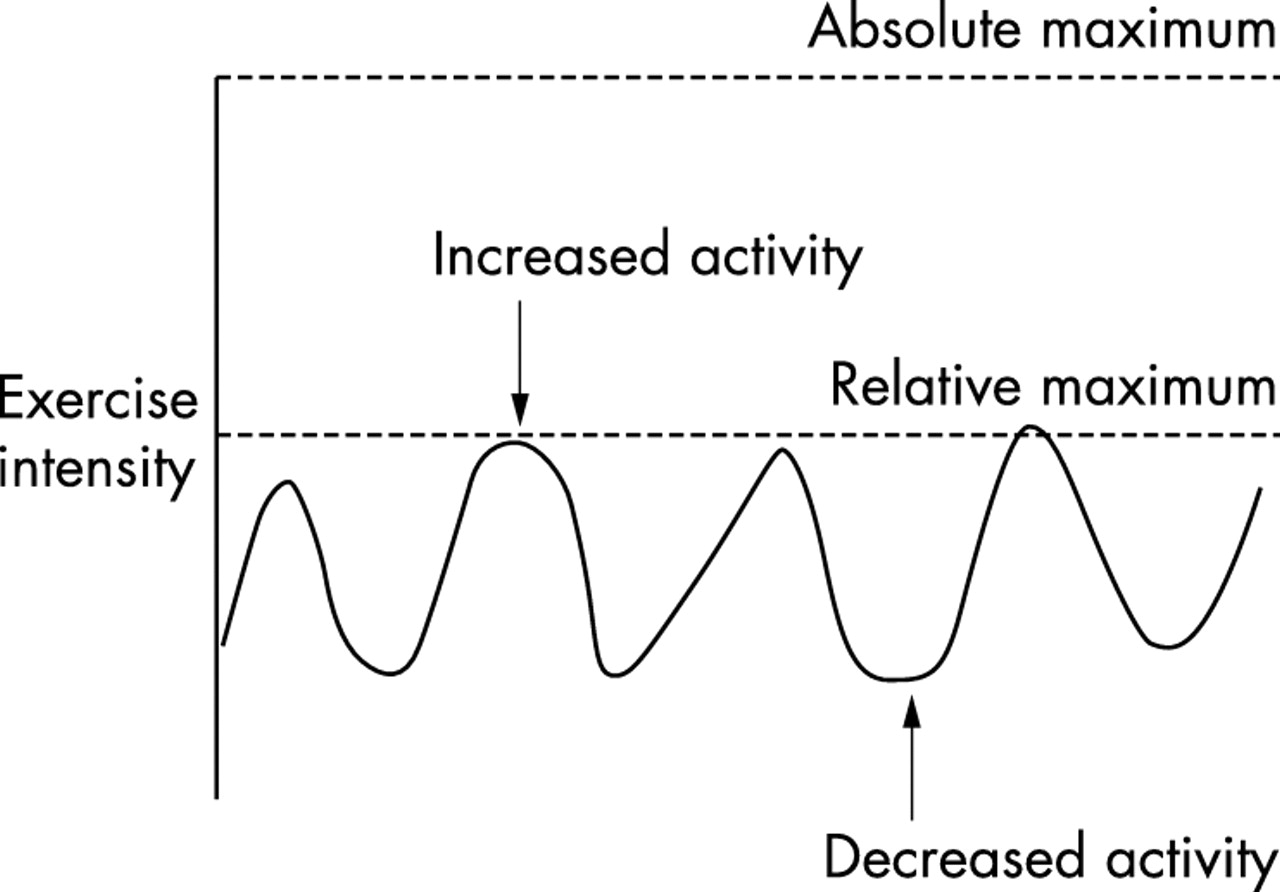
These central mechanisms would reduce efferent neural drive during the fatiguing process. Furthermore, there are three lines of evidence suggesting that all motor units in the exercising skeletal muscle may never be maximally recruited, even at the onset of a maximal isometric contraction.99 The relevance of this finding to the “catastrophe” theory of fatigue was discussed in the companion paper.77
Firstly, muscles artificially stimulated at higher frequencies than are produced during maximal voluntary contractions (MVCs) in vivo produce a greater force output.100 For example, the optimum stimulation rate required to produce a maximum contraction of skeletal muscle is 50–120 Hz, whereas the maximum frequencies measured during MVCs is ∼29 Hz in the deltoid,32 ∼23 Hz in the biceps brachii,101 and ∼11 Hz in the soleus muscle.102 However, a peak frequency of 100 Hz has been measured in single motor units in the adductor pollicis muscle.103 The findings that the maximum in vivo neural frequencies are less than the artificial stimulation rates that produce a maximal force led Enoka100 to conclude that the force achieved during maximum voluntary contractions cannot be the true maximum force. This conclusion is supported by dynamic magnetic resonance imaging techniques, which show that only ∼70% of the available motor units are recruited during a maximal voluntary isometric contraction.104
Secondly, even electrical stimulation may not reveal the absolute maximum force output of which a muscle is capable, particularly during dynamic activity.105 It is unsafe to increase the electrical current beyond a certain strength when attempting to measure contractile force because of the possibility that muscle damage or tendon rupture will result.
Thirdly, maximal isometric and concentric contractions produce less force from more muscle fibres than do eccentric contractions.106,107 Therefore, when muscles contract voluntarily, maximally and isometrically, the force output cannot be maximal because the same absolute quantity of muscle can generate more force when contracting eccentrically.106 It is not yet clear why the same muscle produces different “maximal” force outputs during isometric, concentric, and eccentric contractions. The most likely explanation is that different neural mechanisms are active during these different types of muscle contraction.108
These examples suggest that a degree of muscle recruitment reserve exists during supposedly MVCs and implies that voluntary control is unable to produce an absolutely maximal motor unit recruitment.38,99 However, there are more obvious examples of the complexity of the relation between the generation of force output, fatigue, and perception of effort. For example, when subjects are not encouraged during repeated maximal contractions, their force output is less than when sustained vocal encouragement is provided throughout the exercise.38 Similarly, verbal encouragement increases the duration for which a contraction can be sustained. Therefore, brain areas involved in motivation and effort perception must also play a role in setting “maximal” force output under different environmental conditions and in persons with different diseases.109–113
The reason for this recruitment reserve may be to prevent damage to muscle fibres and other organs, for example the heart,34,114 which could result from excessive muscular activity.
CENTRAL FATIGUE AND ACTIVE CONTROL OF NEURAL DRIVE
Most research on the effect of brain regulation on muscular fatigue has been on isometric contractions or single muscle groups. However, many muscles are active during exercise and all act in a coordinated manner. Furthermore, a problem specific to prolonged dynamic exercise is that the power output increases and decreases in an apparently random manner.115–117 To obviate this problem of dynamic changes in power output during exercise, some researchers have measured force output and EMG activity during an MVC at the beginning and end of exercise on the assumption that changes during the MVC will reflect what transpired during the exercise bout.118 For example, Nicol et al119 reported that the force output and the integrated EMG (IEMG) activity during an MVC decreased by ∼30% in subjects studied before and after a 42 km marathon foot race. They speculated that these changes were caused by either insufficient conscious effort or altered subconscious neural drive.120 Similarly, Sacco et al121 found that prolonged lower limb exercise reduced neural drive to synergistic muscles not directly involved in the exercise.
However, these models permit only a “snapshot” of the neural changes that occur during exercise.122,123 Indeed, it is this “snapshot” approach that may have encouraged the adoption of the linear physiological models (fig 1), as these require fewer measurements than non-linear models.
By adding repeated high intensity 1 and 4 km sprint bouts during a 100 km cycling time trial, a protocol was developed that would allow a “moving picture” rather than a “snapshot” approach.124 This protocol allowed the measurement of “maximal” force output and neural recruitment patterns during different sections of the time trial in which subjects were instructed to complete each 1 and 4 km sprint as fast as possible in the context of completing the entire 100 km cycle, also in the fastest possible time.
Even though the subjects were verbally encouraged to perform at maximal effort, their average power outputs decreased progressively during the consecutive 1 km sprints.111 IEMG activity declined in parallel with these reductions in power output (fig 4), indicating the central brain recruitment of a progressively smaller number of motor units. These changes occurred even though 20% or less of the available motor units in the lower limb were recruited. Heart rate was near maximal during each of the sprints, probably indicating that the subjects consciously attempted to produce a maximal effort, even though the extent of their skeletal muscle recruitment declined progressively.

Mean power output and integrated electromyographic (IEMG) activity during 1 km sprints that occurred during a 100 km cycling time trial. Note that power output and IEMG activity fall in parallel, indicating that the progressive reduction in power output is caused by the recruitment of fewer motor neurones by the brain, hence central fatigue according to fig 2 (lower two lines). *p<0.05, **p<0.01 compared with sprint 1. Data from St Clair Gibson.111
Muscle biopsies performed before and after the trial showed that muscle glycogen concentrations decreased by ∼80% during exercise, even though only 20% or less of the available muscle fibres were recruited at any instant during the trial. If the same 20% of muscle fibres had been recruited continuously for the entire duration of the trial, there would have been complete muscle glycogen depletion only in that 20% with the possible development of muscle rigor. Therefore, either muscle recruitment strategies for the entire limb were altered during the trial or different muscle fibres in the same muscle group were recruited at different times during exercise, with fibres that were initially recruited being replaced by previously unused muscle fibres at the same time as overall neural drive was decreasing. This process has been described as motor unit substitution/rotation.125
These findings appear to indicate that the central nervous system reduces force output by reducing neural drive to the peripheral muscles and hence reducing the number of motor units recruited during exercise, despite the conscious efforts by the participants to maintain maximal effort. The finding that force output decreased in the presence of a large motor unit “reserve” suggests that this reduced neural drive is part of a protective response by the subconscious brain. This protective function would prevent the terminal metabolic crisis predicted by the linear model (fig 1) in which neural drive increases progressively and is not reduced at exhaustion,38 thereby precipitating a catastrophic failure of homoeostasis.
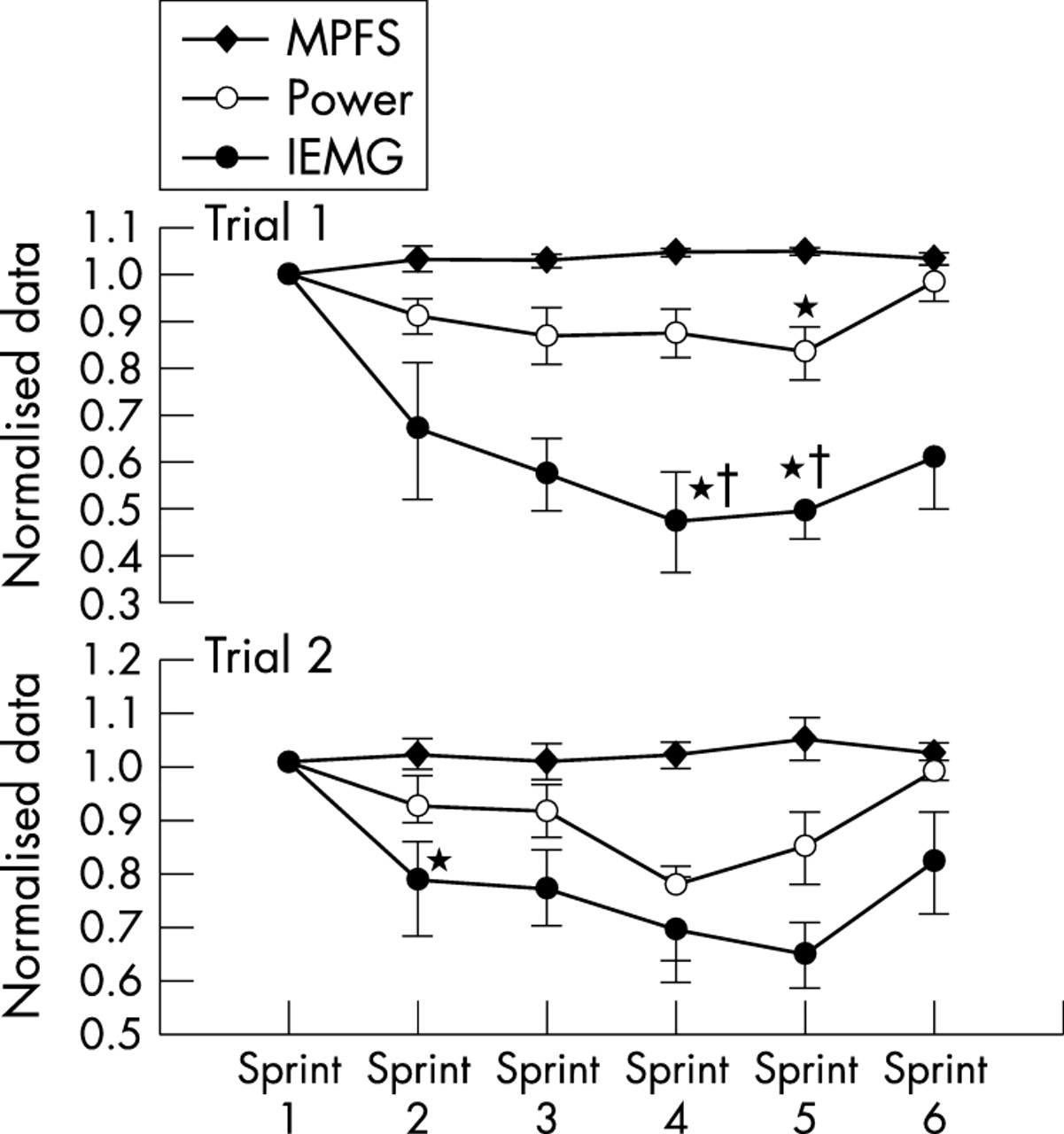
Further studies have supported these interpretations. Kay et al122 modified this original protocol to include six maximal sprints interspersed within a 60 minute cycling time trial, during which power output, IEMG activity, and ratings of perceived exertion were measured (fig 5). There was a reduction in average power outputs and IEMG activity from the second to the fifth sprints. However, both IEMG activity and power output increased during sprint 6, which occurred in the last minute of the ride. This again shows the existence of a subconsciously controlled motor unit recruitment reserve during the initial five sprints, with the ability to recruit that muscle reserve during the final sprint “end spurt”. If substrate depletion or metabolite accumulation caused the decrease in force output and IEMG activity during sprints 1–5, as is predicted by the linear, peripheral fatigue model, there could not have been any increase in power output during the final sprint. As IEMG activity tracked changes in power output, the initial reduction in IEMG activity during sprints 1–5, reversed in sprint 6, could not be explained by changes in temperature, conductivity, or IEMG electrode placement during the trial. Therefore the decrements in IEMG activity during both trials were a real phenomenon and were not an artefact of the testing methods. Indeed it has been previously shown that IEMG activity measured during cycling is both reliable and reproducible.126,127
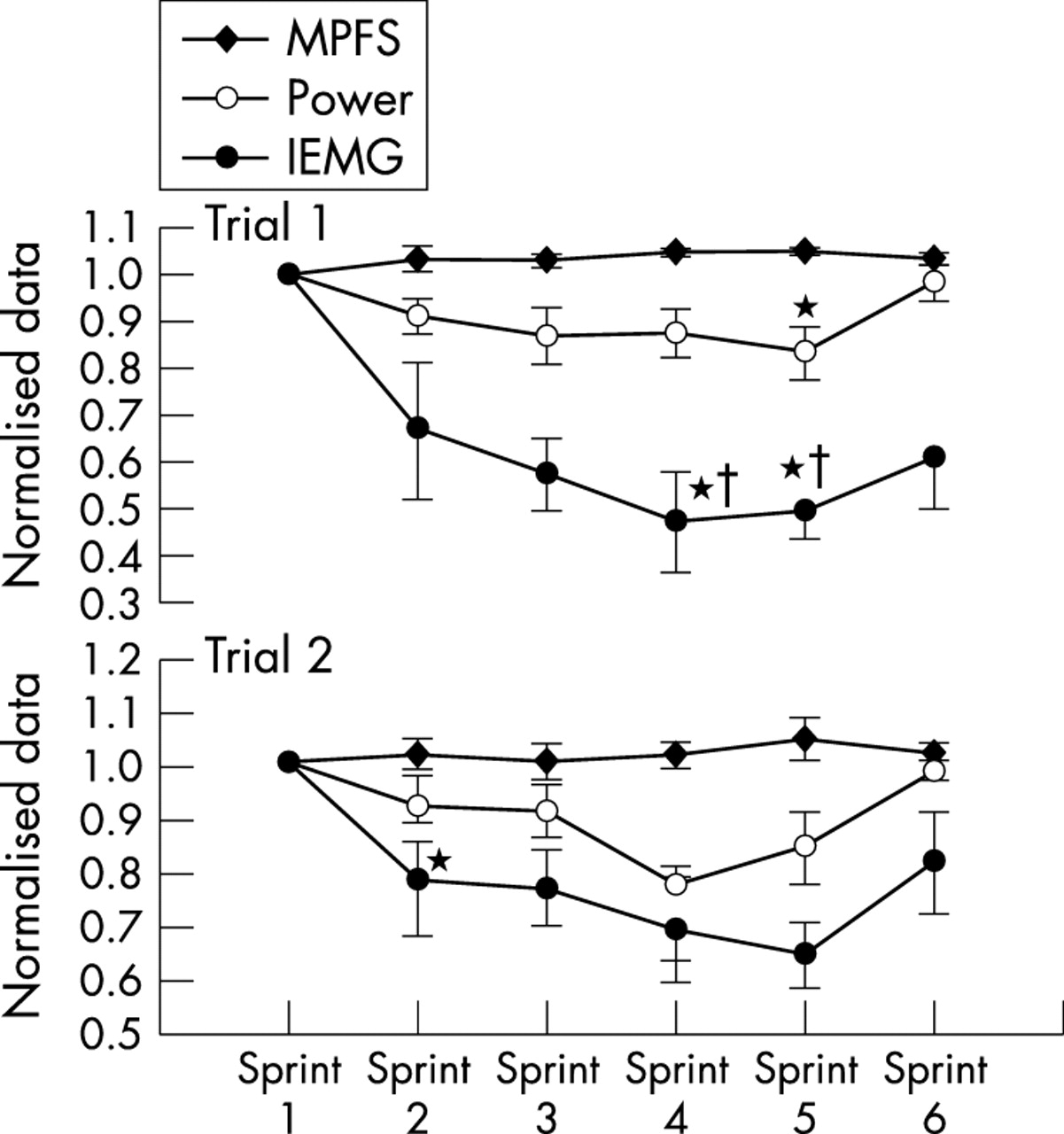
Mean power output and integrated electromyographic (IEMG) activity during one minute sprints during a 60 minute cycling time trial. Note that in both the first and second trials, power and IEMG activity fall in sprints 2–5 but recover in the final sprint. The initial reduction in IEMG activity with recovery in the final sprint indicates the action of a central regulator of power output according to a subconsciously determined pacing strategy. MPFS, mean percentile frequency shift. *p<0.05 compared with sprint 1. †p<0.05 compared with sprint 2. Data from Kay et al.122
A further finding in that study was that the rating of perceived exertion during the sprints, measured on the Borg (0–20) scale, was submaximal and ranged from ∼14 for the first sprint to ∼18 for the last sprint.42 This indicates that despite encouragement to exert themselves “as hard as possible” during each sprint, the subjects maintained a reserve capacity, presumably so that they could complete the entire 60 minute time trial without either physical harm or a catastrophic reduction in pace. As the rating of perceived exertion is probably the overall conscious perception or “gestalt” of all the physiological activity in the body, this finding indicates that regulation must occur at a subconscious level. The internal controls that set the exercise intensity and therefore the pacing strategy must have overridden the conscious desire of the subject, and the explicit instructions given to the subject, to exercise as hard as possible. Indeed, subjects achieved a rating of perceived exertion of only ∼18 during the final sprint even though they knew that the trial was essentially over. This suggests the presence of an overall pacing strategy during exercise, which may also indicate that the activities will also occur after termination of the specific exercise bout. The teleological value of such a control would be obvious. Our evolutionary ancestors, at risk of predation, would not have survived if any bout of physical activity could produce such complete exhaustion that escape from further predatory attack would have been impossible.
CENTRAL INTEGRATIVE CONTROL OF THE EXERCISE INTENSITY: THE CENTRAL GOVERNOR MODEL AND THE ROLE OF FATIGUE SENSATIONS
Integrating these new findings, we therefore propose a new theory of how exercise intensity is regulated: the central integrative, or central governor model (fig 6). In this model, the state of physical exhaustion is considered to be a relative rather than an absolute event. We suggest that, before and continuously during exercise, the brain performs subconscious calculations of the metabolic cost required to complete a given exercise task, and then computes how this will be influenced by the prevailing environmental conditions and the current physical state. This allows the selection of an optimum pacing strategy that will allow completion of the task in the most efficient way while maintaining internal homoeostasis and a metabolic and physiological reserve capacity.
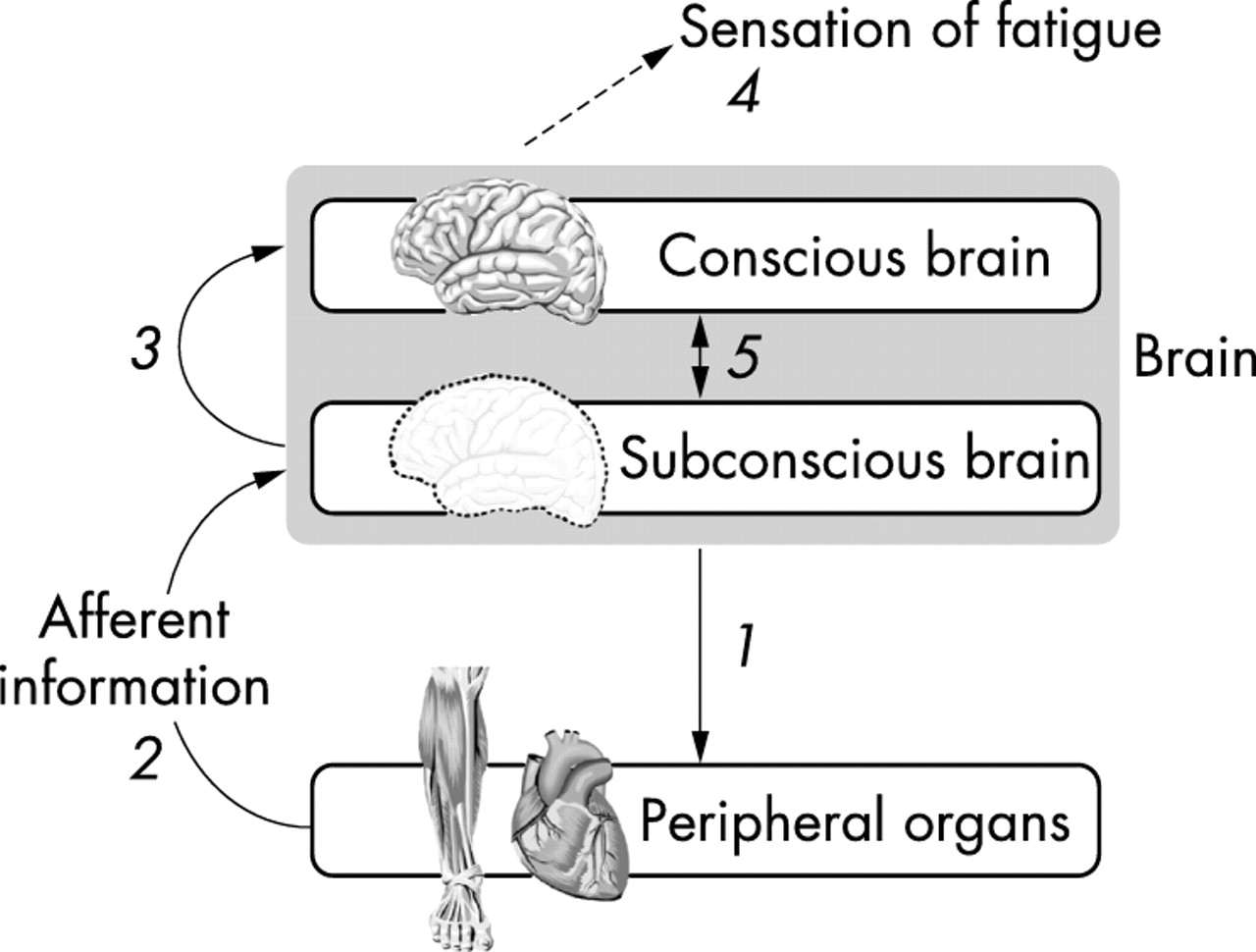
The central integrative model of exercise regulation. This model proposes that the subconscious brain sets the exercise intensity by determining the number of motor units that are activated and hence the mass of skeletal muscle that is recruited throughout the exercise bout (1). The extent of motor neurone activity and hence skeletal muscle motor unit recruitment can then be influenced by sensory feedback from a variety of peripheral organs (2), yet to be characterised and which probably include the skeletal muscles, the respiratory muscles, the heart, and central and peripheral chemoreceptors and mechanoreceptors. From the beginning of the exercise bout, the subconscious brain informs the conscious brain (3) of an increasing neural effort, perhaps related to an increased difficulty in maintaining homoeostasis at that exercise intensity, and this is interpreted by the brain as the increased sensation of fatigue (4), which may itself control further subconscious brain control processes (5).
This theory is an extension of that originally proposed by Ulmer128 who defined this “resetting” of power output or speed as “teleoanticipation”. He suggested that a central “programmer”, probably working at a subconscious level, calculates the time necessary to complete an activity and include this in a calculation of the required power output to complete the task while maintaining homoeostasis in all the bodily systems.
The subconscious mental calculations that form the basis of the central integrative model would therefore act to regulate the power output during all forms of exercise as part of the controls necessary to maintain whole body homoeostasis. The conscious sensation of fatigue would therefore be one manifestation of these subconscious mental calculations and the resultant alterations in neural drive (fig 6). Alternatively, the sensations of fatigue may represent the calculated neural “effort” expended in maintaining whole body homoeostasis so that the progressive increase in these sensations during exercise indicates how close the subject is to the end of an exercise bout.
Thus, in this model, the symptom of fatigue may have a cognitive function, in which the subconscious processes of the brain that are responsible for planning and regulating the physical activity, project also to the conscious brain, creating the sensory perception of fatigue. Fatigue symptoms so generated would modify or inhibit the conscious desire to work harder than the subconsciously planned strategy, in response for example, to distracting sensory input such as vocal support from spectators. If such vocal support were able significantly to alter the subconsciously derived pacing strategy, homoeostasis would be threatened. Therefore, this model predicts a novel interpretation of the meaning of fatigue. For it suggests that fatigue is a symptom and not a physical state. By indicating the extent of the bodily effort already activated to maintain homoeostasis at that chosen exercise intensity and therefore the extent of the reserve that could support any further increase in work output, the symptom of fatigue acts as the additional conscious “restrainer” or modulator of the exercise intensity.
Perhaps the most important input to the calculation made by the central integrator is the known duration of the activity. When either the distance or time required to complete the activity is known before the exercise begins, the activity is defined as a “closed loop” activity.99 In contrast, if either the distance or duration is unknown beforehand, so that there is no fixed end point to the exercise, it is defined as “open loop” activity. The central integrative strategies would be very different for closed and open loop activity, because of the absence of a known end point in open loop activity so that the necessary calculations cannot be computed by the brain before or during exercise. The ability of subjects to faithfully reproduce almost identical pacing strategies and hence overall performances when completing closed loop exercise tests in the laboratory, even when they receive minimal external information,129,130 is perhaps the most striking evidence for the presence and power of this central integrative mechanism. In contrast, performance during open loop exercise is notoriously variable.
The presence of such a central integrative mechanism is, however, not unique to humans. For example, ants can gauge with superb accuracy, the distances they travel over a range of different terrains.131,132 This ability was more closely related to proprioceptive input from different body segments than to metabolic rate or visual clues. Furthermore, before migration, birds successfully calculate the metabolic requirements of their flight and increase the quantity and alter the composition of their fuel stores, embarking on their migration only when they have sufficient body fat stores.133 They also modulate their flight speed and flying patterns during the migratory flight to accommodate the extra body weight caused by their increased fuel reserves. It is also interesting that entire flocks of birds migrate as a unit. Therefore their pacing strategies appear to be either universal or commonly linked.128
COMPLEX SYSTEM ACTIVITY AND THE CENTRAL GOVERNOR THEORY OF FATIGUE
The central governor model conceives the physical state of exhaustion, as opposed to the sensory perception of fatigue, as a relative rather than an absolute condition. Skeletal muscle work is not determined by absolute metabolic changes in the active muscles but, rather, is continuously reset, decreasing or increasing as a result of continuous subconscious integrative calculations with the end point of the exercise acting as the reference point. In this model, the development of the physical component of fatigue becomes a relative process as the exercise intensity is constantly changed during exercise,33,122 with the central governor either increasing motor unit recruitment or derecruitment throughout the exercise bout as a result of these calculations.
The study of Palmer et al115 provides evidence for this continuous, integrative process. They measured continuously the heart rates of four subjects during a 104 km cycling race. Heart rates, which generally represent exercise intensity,44,134 increased and decreased in an apparently random manner in all four cyclists continuously throughout the event (fig 7A). The changes in heart rate were not related solely to geographical changes during the race (fig 7B) as these changes also occurred when the cyclists were progressing over relatively flat sections of the course (race distances 20–45 km; fig 7B). These findings have been confirmed in other studies of more experienced, professional cyclists competing in the Vuelta a Espana three week cycle race.117

Individual heart rates of five cyclists (S1–S5) during a 104 km cycling race. Note the oscillatory nature of the individual heart rates (A) even during that section of the race completed on the flatter section of the course (20–45 km; B). Data from Palmer et al.115
We suggest that these variable changes in heart rate are an example of a non-linear dynamic process. In non-linear dynamics, the physiological changes occur as part of a complex system integration, in which many physiological changes interact with each other through different feedback and feedforward systems. These complex interactions create alternating changes in the different physiological variables,135–137 with these oscillatory changes depending on the complexity of the regulatory system.
Complex system theory developed as a scientific discipline in the 1960s and 1970s138 to counter the trend of increasing specialisation in individual systems or organs. This specialisation led to the reductionist scientific approach grounded on the belief that complicated systems could be analysed by reducing them to simpler subdivisions and analysing each part separately.139,140 Scientific disciplines such as biochemistry, genetics, and molecular biology have developed using this reductionist approach, and are now regarded as separate scientific disciplines. Each of these disciplines searches for different components of physical structures or activity that may be responsible for controlling human physiology and anatomy. Although this reductionist approach has produced important scientific breakthroughs, it has become increasingly evident that individual variables studied separately cannot explain the complex behaviour of human metabolism during exercise.139 This insight stimulated the study of how different systems interact to create this complex behaviour.
Complex system theory examines the interrelatedness of individual physiological processes, in the belief that the interaction of these different processes is as important as the function of any one of the individual processes. Complex system analysis has been used to assess relations between systems and structures in such diverse processes as electrical power grids, food webs, neural networks, internet connections, and cellular and metabolic networks.137,141 It has been used to examine the generation of biological rhythms such as heart rate rhythms in both healthy and diseased subjects, the motion of upper and lower limbs during walking and running, and hormonal cycles in which the pulsatile release of hormones is necessary for the regulation of metabolism and multiorgan function.135
A central concept in complex system theory is that different variables affect each other constantly either directly or in a feedforward or feedback manner. Complex system theory also shows the importance of different levels of interaction, such that physiological activity at the molecular level is dependent on cellular changes, which are dependent on anatomical organ changes, which in turn depend on whole body changes, changes in the human/environment interface, changes in society, and finally changes in the entire world system. In this model, there is a two way interaction, with change at “lower” levels also affecting activity at “higher” levels.139 The complex system model also suggests that different physiological activities such as electrical and magnetic activity and protein and genetic structures, all of which occur in different body tissues or functional processes, also interact and affect each other.
An example of how different variables affect each other can be found in the description of complex system interaction involving the effect of gravity on the trajectory of different numbers of planets in a solar system.142 It is relatively easy to calculate the trajectory of a smaller planet around a larger planet if the gravitational force of both planets is known and no other planets are present in the system. Furthermore, the position and trajectory of the smaller planet could also be relatively easily calculated from knowledge of its current position. However, if a third planet is introduced to the system, the gravitational pull of the third planet affects the trajectories of the two other planets. This triple interaction creates a more random appearing pattern to the trajectories of all three planets. The level of mathematical calculation required to determine the future position and trajectory of any one planet becomes almost impossibly difficult, as the gravitational fields of each planet affects the other planets differently and continuously. If fourth and fifth planets are added to the system, the trajectories of each planet and positions relative to each other would appear increasingly haphazard, and the planets would appear to move in a completely random manner. But, underlying this apparent randomness would be deterministic interactions between the different gravitational forces of each planet acting collectively.
Therefore, in a complex system, any observed change in one variable at a particular moment is the result of changes in the interaction of that variable with other variables in the system at some earlier moment. Furthermore, that variable may appear to be changing in a random manner. However, as suggested by Gleick,138 the interaction of the other variables is contained in the changing history of the variable being observed.
A complex non-linear dynamic system therefore requires a deterministic control mechanism and a fluctuating environment in order to generate alternating or oscillatory changes in physiological variables.135 During exercise the desire to reach the end point in the fastest possible time would be the deterministic control, and the fluctuating environment would be provided by changes in both the external environment and the internal physiological milieu.
Therefore, according to a central governor model which incorporates complex system dynamics, alterations in exercise intensity would occur in response to metabolic changes in the periphery and would be orchestrated by central integrative controls. For example, if blood glucose concentrations decreased, exercise intensity would decrease similarly. This decrease in exercise intensity would reduce peripheral glucose use allowing the blood glucose concentrations to increase, thereby allowing the exercise intensity also to increase (fig 8). The model becomes a complex system when inputs from additional variables such as rates of muscle glycogenolysis and lactate production, cardiovascular and respiratory variables, and rates of heat production and storage are also included. The end result of all these changes, integrated by the central governor and regulated by motor unit recruitment or derecruitment would produce the fluctuating alterations in skeletal muscle force output that appears to be a defining characteristic of human exercise performance. Thus this non-linear complex system model provides a more complete description of homoeostatic control, in which changes in any system cause a response, the aim of which is to oppose these changes as soon as possible as part of a protective mechanism. This function of the complex non-linear model is discussed further in the companion paper by Lambert et al.143

Non-linear oscillations in exercise intensity could be caused by oscillations in the number of motor units that are allowed to be recruited by the central integrator in the subconscious brain as a consequence of calculations made on the basis of sensory inputs from multiple peripheral and central sensors. The goal of the central integrator is to maintain homoeostasis at a level of neural effort, sensed as the perception of fatigue, that the individual athlete will accept.
This model can then be used to describe the changes described by Kay et al122 (fig 5). The initial pace chosen in the first interval was subconsciously calculated on the basis of prior experience. However, as the trial progressed, metabolic and other changes would have initiated subconscious integrative calculations which would establish that the initial pace would require too great a neural effort, and hence too strong sensations of fatigue, to maintain homoeostasis at that exercise intensity. As a result of these subconscious calculations, power output was decreased in the following sprints. However, as the last sprint approached, these calculations would establish that the intensity could be increased for the final minute.
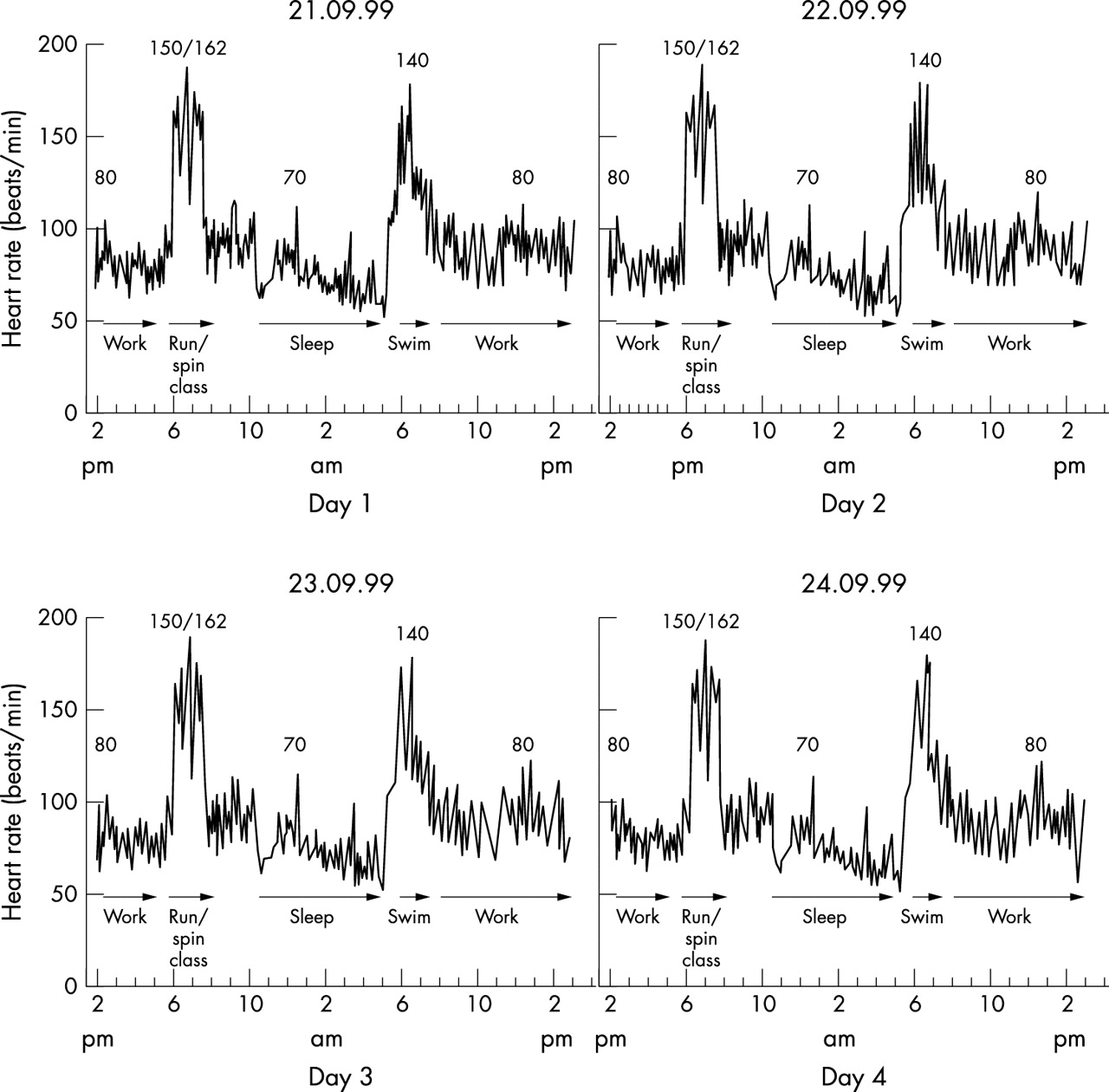
This pattern of altering power output during exercise is also part of a macro cycle which occurs both during and after each exercise bout. For example, each single exercise bout becomes part of a longer, lower frequency oscillation in which a number of days of consecutive activity are performed (fig 9). On an even larger scale, this oscillation in physical activity would alter in response to seasonal variation or the aging process as part of the habitual lifestyle. Thus, according to this complex model, death is the only absolute end point of “exercise” activity.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Heart rates measured for four consecutive days in a female athlete. The heart rate pattern is similar during both exercise and activities of daily living for each of the four days (M Lambert et al, unpublished data).
In summary, we propose that skeletal muscle power output during exercise is regulated by a governor mechanism in the brain as part of a complex integrative system. This contrasts with the more usually described linear model in which it is assumed that a single variable regulates the entire system so that the maximal capacity of the system is determined by the maximal activity of that particular variable. However, in reality, multiple variables in multiple physiological systems interact together over time. The continuously altering skeletal muscle power output during exercise is caused by the continuous interaction of each of the different variables in the different physiological systems. The efferent motor command from the central brain region would be both a response to and a controller of these different oscillatory interactions. This function of this complex system interaction would be to maintain homoeostasis. In this system, the different homoeostatic mechanisms interact in order to protect the individual from a terminal metabolic crisis that would force exercise to stop perhaps with the risk of physical damage. The complex integrative governor system would also explain the continuous alterations in heart rate and pacing strategies evident in athletes during competition115,117 and in individuals during their normal activities (fig 9).
CONCLUSIONS
Here we propose that the development of physical exhaustion needs to be understood as a relative rather than an absolute event. We also argue that the sensation of fatigue is simply the sensory representation of the underlying neural integrative processes, rather than a defined and measurable physical event, that is a reduced skeletal muscle force output. We also propose that exercise activity is controlled as part of a pacing strategy involving active neural calculations in a “governor” region of the brain, which integrates internal sensory signals and information from the environment to produce an acceptable exercise intensity that will still allow homoeostasis to be maintained at a level of sensory effort, the “sensation of fatigue”, that the individual athlete finds acceptable. In this system, the end point of the exercise bout is the controlling variable. This process is an example of a complex non-linear dynamic system in which physiological systems interact as part of a complex system to regulate activity before, during, and after the exercise bout.