Article Text

Abstract
Objectives: Sequence variation within the COL5A1 and TNC genes are known to associate with Achilles tendinopathy. The primary aim of this case-control genetic association study was to investigate whether variants within the matrix metalloproteinase 3 (MMP3) gene also contributed to both Achilles tendinopathy and Achilles tendon rupture in a Caucasian population. A secondary aim was to establish whether variants within the MMP3 gene interacted with the COL5A1 rs12722 variant to raise risk of these pathologies.
Methods: 114 subjects with symptoms of Achilles tendon pathology and 98 healthy controls were genotyped for MMP3 variants rs679620, rs591058 and rs650108.
Results: As single markers, significant associations were found between the GG genotype of rs679620 (OR = 2.5, 95% CI 1.2 to 4.90, p = 0.010), the CC genotype of rs591058 (OR = 2.3, 95% CI 1.1 to 4.50, p = 0.023) and the AA genotype of rs650108 (OR = 4.9, 95% CI 1.0 to 24.1, p = 0.043) and risk of Achilles tendinopathy. The ATG haplotype (rs679620, rs591058, and rs650108) was under-represented in the tendinopathy group when compared to the control group (41% vs 53%, p = 0.038). Finally, the G allele of rs679620 and the T allele of COL5A1 rs12722 significantly interacted to raise risk of AT (p = 0.006). No associations were found between any of the MMP3 markers and Achilles tendon rupture.
Conclusion: Variants within the MMP3 gene are associated with Achilles tendinopathy. Furthermore, the MMP3 gene variant rs679620 and the COL5A1 marker rs12722 interact to modify the risk of tendinopathy. These data further support a genetic contribution to a common sports related injury.
Statistics from Altmetric.com
There is a spectrum of pathology that can affect the Achilles tendon and surrounding structures.1 Achilles tendinopathy (AT) is one of these pathologies which is a painful and degenerative condition that affects individuals who participate in a range of sporting pursuits as well as the less physically active.2 3 Acute spontaneous rupture is another common pathology that can affect the Achilles tendon, particularly in the middle-aged, male athlete.4 A number of intrinsic and extrinsic factors have been implicated in raising the risk of both AT and Achilles tendon rupture.5
In recent years evidence has emerged that Achilles tendon pathology has a genetic component. For example, Mokone and colleagues have identified that a GT repeat variant within the gene that encodes the tenascin C protein (TNC), a key constituent of tendon which is regulated by mechanical loading,6 7 is associated with AT and rupture.8 In addition to the TNC gene, polymorphisms within the 3′-untranslated region of the COL5A1 gene have also been associated with AT in both South African9 and more recently an Australian population.10 However, variation within the Sp1 binding site of the COL1A1 gene does not appear to associate with Achilles tendon pathology,11 nor do polymorphisms within the related COL14A1 and COL12A1 genes.12 Definitive biochemical mechanisms that explain these observations are presently unknown and should form the focus of future investigations.
In addition to the TNC and COL5A1 genes, Achilles tendon pathology might also be associated with genes that encode proteins with regulatory roles in maintaining extracellular matrix (ECM) homeostasis. Included in this class are the matrix metalloproteinases (MMPs). The MMPs are known to consist of over 20 distinct endopepdidases that can catalyse a broad spectrum of both ECM and non-ECM substrates.13 One of the family, MMP3, can catalytically degrade multiple substrates including: types II, IV, V, IX, X collagens, laminin, fibronectin, proteoglycan, decorin and aggrecan.13 14 MMP3 is also known to activate, via propeptide removal, several other MMPs.13 15 Expression of the MMP3 gene can be substantially altered by the 5A/6A promoter polymorphism16 and this variant has been associated with a number of pathological states.17 18 Interestingly, reduced levels of MMP3 mRNA19 20 and immunochemically detectable MMP3 protein have been observed in resected AT tissue compared to control tissue.20
Although there has been previous speculation about the possible role of MMP genes in tendinopathy21 22 the hypothesis has, to our knowledge, never been tested. Accordingly, the aim of this investigation was to establish whether variants within the MMP3 gene were associated with AT or Achilles tendon rupture. A secondary aim was to investigate whether MMP3 and COL5A1 gene-gene interactions were involved in these Achilles tendon pathologies.
MATERIALS AND METHODS
Subjects
In all, 114 Caucasian subjects diagnosed with Achilles tendon injuries, including 75 with chronic AT (TEN) using clinical criteria and 39 with partial (n = 3) or complete ruptures of the Achilles tendon (RUP), were recruited for this study from the medical practice at the Sports Science Institute of South Africa and other clinical practices within the greater Cape Town area of South Africa. Rupture of the Achilles tendon was confirmed during surgery or by imaging. Ten of the subjects in the RUP group had a history of tendinopathy. An additional 98, apparently healthy, unrelated, Caucasian subjects without any history of symptomatic Achilles tendon injuries were recruited as controls (CON). The inclusion and exclusion criteria of the participants have been previously described.8 9
Prior to participation in this study, the subjects gave informed written consent and completed medical history questionnaire forms. This study was approved by the Research Ethics Committees of the Faculty of Health Sciences within the University of Cape Town, South Africa and the University of Northampton, England.
DNA extraction and single nucleotide polymorphism (SNP) selection
DNA was extracted using the procedure described by Lahiri and Nurnberger23 and modified by Mokone et al.8
Single nucleotide polymorphisms (SNPs) within the MMP3 gene and its 5′-flanking sequence were identified from databases hosted by the National Centre for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/) and the International HapMap project (http://www.hapmap.org/). The MMP3 gene is transcribed in the negative orientation resulting in the mRNA sequence corresponding to the bottom DNA strand. As indicated in fig 1, the complimentary nucleotides are given in the databases for some of the SNPs. These complementary nucleotides are used in this manuscript to avoid confusion. Seven exonic SNPs were identified and annotated onto a schematic diagram (fig 1). Two of these SNPs were nonsynonomous (SNPs which change the amino acid sequence in the gene product), of which only one, rs679620 (E45K) within exon 2, has a high heterozygous frequency and could therefore be potentially informative in genetic case-control association studies. Two of the five synonomous SNPs, rs602128 (D96D) within exon 2 and rs520540 (A362A) within exon 8, also have a high heterozygous frequency and were therefore considered potentially informative. From the databases hosted by the International HapMap project these three exonic SNPs formed a single haplotype block together with 4 informative (high heterozygosity) intronic SNPs that spanned the entire gene and a single informative SNP (rs645419) at -2 kb within the promoter region (fig 1). The functional 5A/6A (rs3025058, -/T) polymorphism, which is often used in association studies involving the MMP3 gene17 18 is located downstream of SNP rs645419. Four major haplotypes which contained these eight potentially informative SNPs were identified using HaploView V.4.124 and the HapMap CEU data (release 22). The same four haplotypes with similar frequencies were also identified when only three of the eights SNPs were analysed. In this study, the MMP3 gene was therefore genotyped for SNPs (1) rs679620 a A/G transition at nucleotide position 28 within exon 2, E45K; (2) rs591058 a T/C transition at nucleotide position 1547 within intron 4, and (3) rs650108 a G/A transition at position 495 within intron 8.
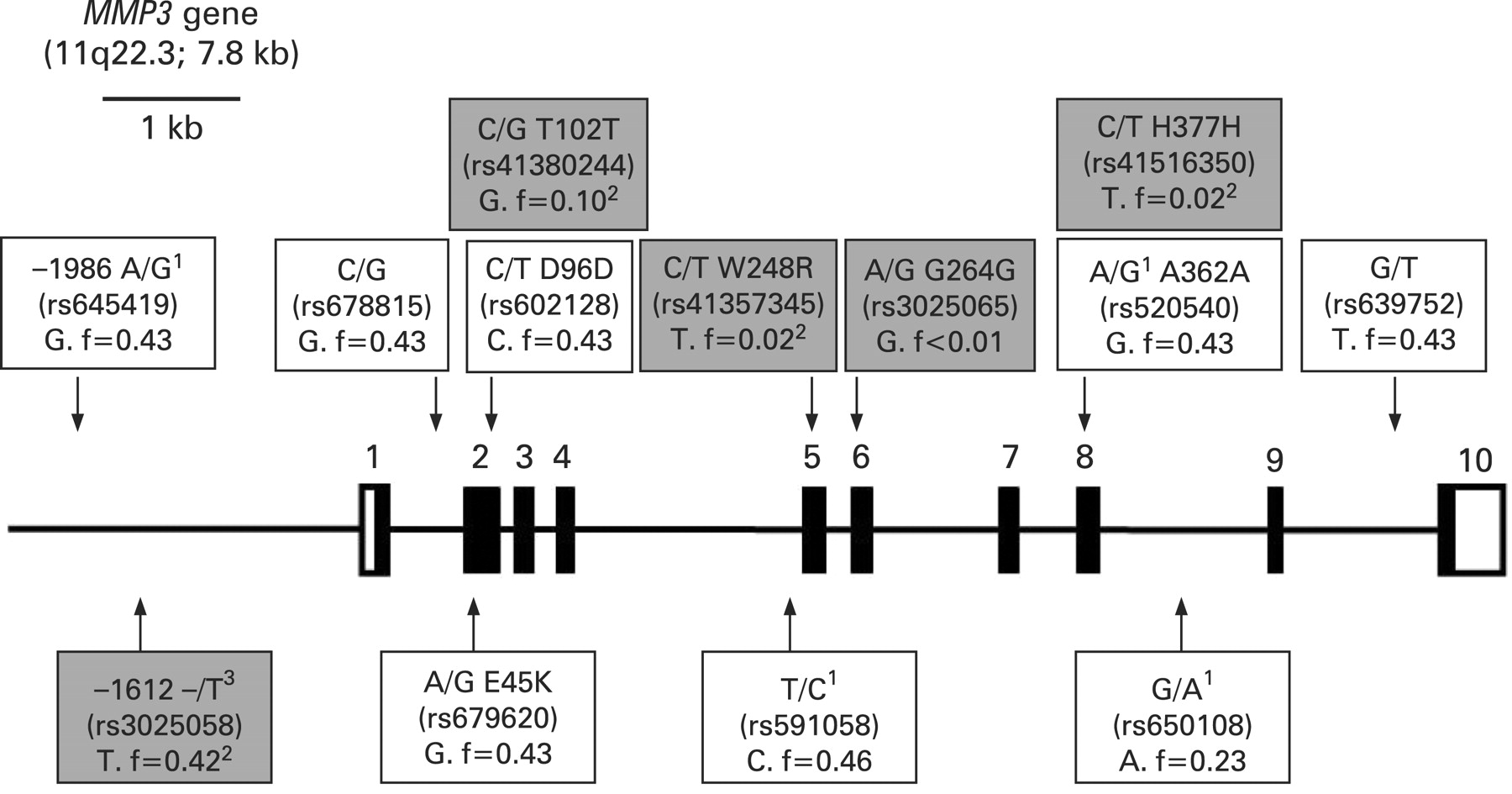
A schematic representation of the exon (rectangles) and intron (horizontal lines) boundaries of the MMP3 (matrix metallopeptidase 3) gene, which is located in the negative orientation on chromosome 11. The translated regions of the exons are solid while the untranslated regions (UTRs) are clear. Exon numbers are indicated. The chromosomal location and the size of the gene are given in brackets. Single nucleotide polymorphisms (SNPs) with high heterozygous frequencies (>20%) identified from databases hosted by the international HapMap project are annotated (clear boxes). In addition, three synonomous, one nonsynonomous and a promoter (rs3025058, -/T) SNPs identified from databases hosted by the National Centre for Biotechnology Information (NCBI) are also annotated (grey boxes). The three SNPs used in this study are annotated below the gene as clear boxes. Accession numbers and base changes are indicated for all the SNPs. The minor alleles and, unless otherwise indicated, their frequencies from the HapMap CEU population are also indicated. Where applicable the amino acid change and number of the exonic SNPs, as well as, the nucleotide position of the two SNPs within the promoter region are indicated. All the information used to construct this figure was obtained from databases hosted by the NCBI and Ensembl (http://www.ensembl.org/index.html) as well as from Beyzade et al.16 1The complementary nucleotides are given in the databases and used in this manuscript to avoid confusion. 2Since frequency data was not available for the HapMap CEU population, minor allele frequency data from other available European population data within the NCBI databases are indicated. 3Also referred to as the -1171 5A/6A polymorphism.
MMP3 SNP genotyping
DNA samples were genotyped for all three MMP3 variants using fluorescence-based Taqman technology (Applied Biosystems, Foster City, CA, USA). Allele specific probes and flanking primer sets (table 1) were used along with a pre-made PCR mastermix containing ampliTaq DNA polymerase Gold (Applied Biosystems, Foster City, CA, USA) in a reaction volume of 25 μl. PCR consisted of a 10 minute heat activation step (95°C) followed by 40 cycles of 15 s at 92°C and 1 minute at 60°C. PCR was performed on an MJ Miniopticon thermocycler (BioRad Laboratories Inc, Herts, UK) and genotypes were determined by endpoint fluorescence using MJ Monitor analysis software (V.3.1).
Statistical analyses
Assuming an allele frequency of 0.43 (observed for MMP3) in the CON group, group sizes of 97 in each group would be adequate to detect an allelic odds ratio of at least 2.25 at a power of 80% and significance level of 5%. Linear and logistic regression models were used to assess difference between the characteristics of the TEN, RUP and CON groups for quantitative and categorical data, respectively. Logistic regression was used to compare the combined, as well as the separate (TEN and RUP) Achilles tendon cases to the control groups, with respect to genotype, allele and haplotype frequencies. Two different methods were used to assess gene-gene interaction between MMP3 and COL5A1 (SNP rs12722). Allele combinations consisting of the markers on the two different genes were constructed and their association with case-control status was tested. The odds ratio based multifactor dimensionality reduction (ORMDR) method25 was also used to select from all SNPs, the pair with the strongest association with tendinopathy. An odds ratio (OR) is the quantitative measure of disease risk being analysed but this is the estimated relative risk of disease with a specific combination of genotypes. Data was analysed using the freely available programming language R (http://www.r-project.org/) and specifically packages DGC-genetics (LD, Hardy-Weinberg, genotype and allelic association), haplo.stats (inferred haplotype association) and ORMDR (interaction between loci on case-control status). Pass2008 (http://www.ncss.com/) was used for sample size calculation.
RESULTS
Subject characteristics
The TEN, RUP and CON groups were similarly matched for age, height, gender and country of birth (table 2). The age of the TEN and RUP groups was the age of initial onset of symptoms of Achilles tendon injury, which were on average 7.8 (SD 8.0) and 7.5 (SD 8.9) years prior to recruitment in this study respectively. The reported weights were also at recruitment, not at time of onset of injury. The TEN and RUP groups were on average significantly heavier with corresponding higher BMI than the CON group. In addition the RUP group was also on average significantly heavier with corresponding higher BMI than the TEN group. There were no MMP3 single nucleotide polymorphism genotype effects on any of the subject characteristics (results not shown).
Twenty six (38.2%) and five (13.5%) of the subjects were diagnosed with bilateral chronic AT and Achilles rupture respectively. Multiple Achilles tendon injuries (greater than one) were documented in 16 (22.9%) and 13 (35.1%) of the TEN and RUP subjects respectively. Forty four percent of the TEN and 35% of the RUP subjects reported either a bilateral and/or multiple Achilles tendon injury.
Genotype and allele frequencies
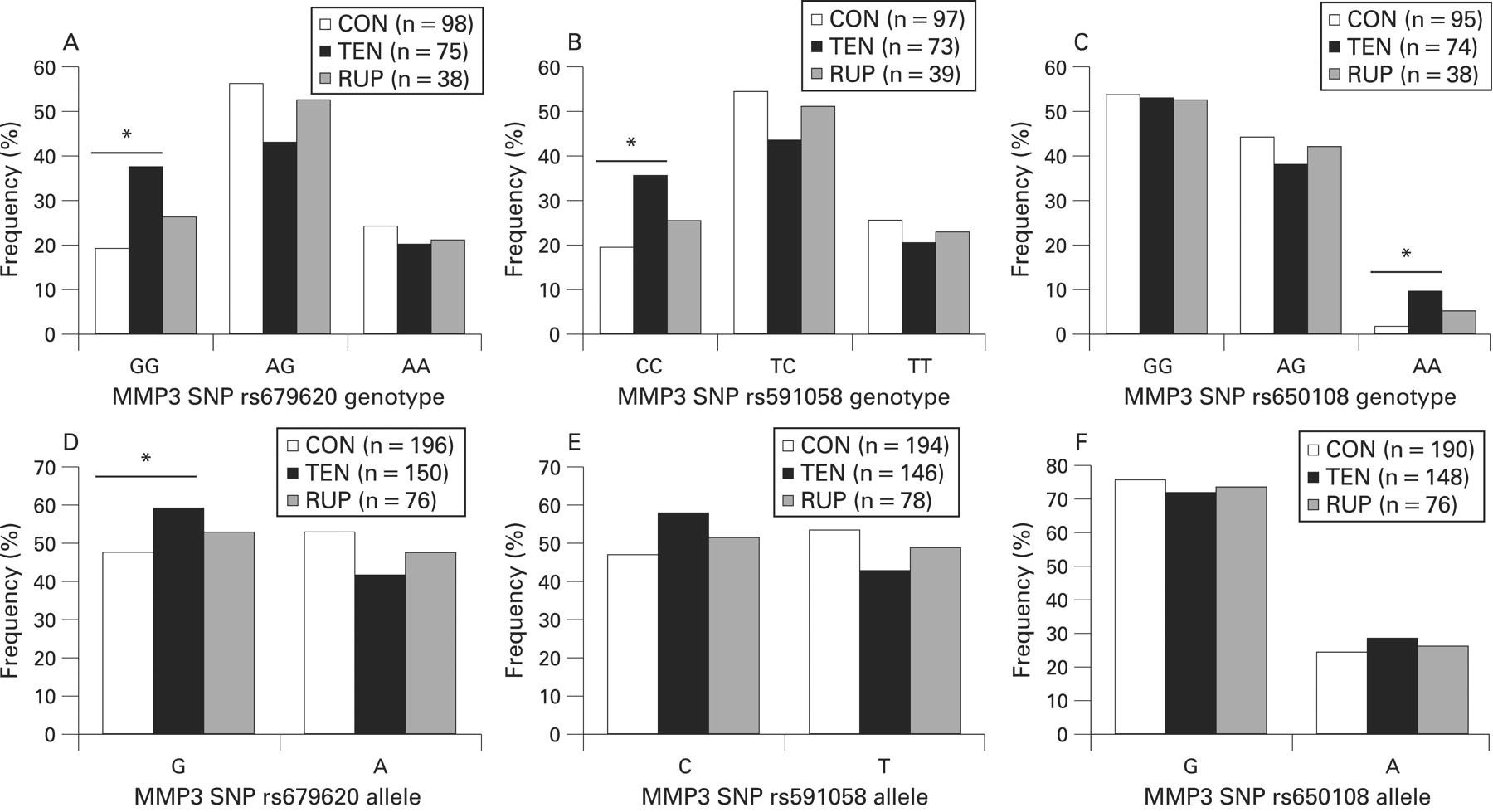
There were no significant differences in the genotype distributions of SNPs rs679620 (p = 0.064), rs591058 (p = 0.117) and rs650108 (p = 0.132) between the Achilles tendon pathology (combined TEN and RUP) and control groups (see figs 2A–C). Similarly, there were no significant differences in the allele distributions of SNPs rs679620 (p = 0.057), rs591058 (p = 0.083) and rs650108 (p = 0.404) between the combined pathology and control groups (figs 2D–F). Since differences have been detected in genotype distributions between subjects with chronic AT or Achilles tendon ruptures,12 the Achilles tendon pathology group was sub-divided into tendinopathy (TEN) and rupture (RUP) sub-groups. There were significant differences in the distribution of the genotype (p = 0.031) (fig 2A) and allele (p = 0.037) (fig 2D) frequencies of the MMP3 rs679620 SNP between the CON and TEN groups. The GG genotype was significantly over-represented in TEN group (37.3%, n = 28) when compared to the CON group (19.4%, n = 19) (p = 0.010, OR = 2.5, 95% CI 1.2 to 4.9). The differences in the genotype (figs 2B and 2C) or allele (figs 2E and 2F) frequency distributions of the MMP3 rs591058 (genotype p = 0.065 and allele p = 0.051) and rs650108 (genotype p = 0.093 and allele p = 0.368) between the CON and TEN groups were not significant. But the CC genotype of SNP rs591058 was over-represented in the TEN (35.6%, n = 26) compared to the CON (19.6%, n = 19) (p = 0.023, OR = 2.3, 95% CI 1.1 to 4.5) and the AA genotype of SNP rs650108 was over-represented in the TEN (9.5%, n = 7) compared to the CON (2.1%, n = 2) (p = 0.043, OR = 4.9, 95% CI 1.0 to 24.1). Similar results were obtained when the 10 RUP subjects with a history of tendinopathy were included in the analysis as part of the TEN group (results not shown).

Relative genotype (A–C) and allele (D–F) frequencies of the MMP3 single nucleotide polymorphisms (SNPs) rs679630 (A and D), rs591058 (B and E) and rs650108 (C and F) for the asymptomatic control (CON, clear bars), chronic AT (TEN, solid bars) and Achilles rupture (RUP, hatched bars) groups. (A) rs679620: TEN vs CON, p = 0.031; RUP vs CON, p = 0.666; (B) rs591058: TEN vs CON, p = 0.065; RUP vs CON, p = 0.734. (C) rs650108: TEN vs CON, p = 0.093; RUP vs CON, p = 0.627. (D) rs679620: TEN vs CON, p = 0.037; RUP vs CON P = 0.500. (E) rs591058: TEN vs CON, P = 0.051; RUP vs CON, p = 0.592. (F) rs650108: TEN vs CON, p = 0.368; RUP vs CON, p = 0.754. The asterisks and solid lines represent specific genotype (A) GG, p = 0.010; (B) CC, p = 0.023; (C) AA, p = 0.043; or allele (D) G, p = 0.037; significant differences between the TEN and CON groups. The number (n) of subjects (A to C) or alleles (D to F) are in parenthesis. CON, control group; RUP, Achilles tendon ruptures; TEN, tendinopathy.
There were, however, no significant differences in the distribution of the genotype (rs679620, p = 0.666; rs591058, p = 0.734; and rs650108, p = 0.627) (figs 2A, 2B and 2C) and allele (rs679620, p = 0.527; rs591058, p = 0.604; and rs650108, p = 0.840) (figs 2D, 2E, 2F) frequencies of the three MMP3 SNPs between the CON and RUP groups. The three MMP3 SNP genotype distributions within the CON, TEN and RUP groups were in Hardy-Weinberg equilibrium. Although there was a reduction in statistical power, similar genotype distributions were nevertheless observed when only the South African born subjects were analysed (results not shown).
Linkage disequilibrium and inferred haplotype analysis of MMP3
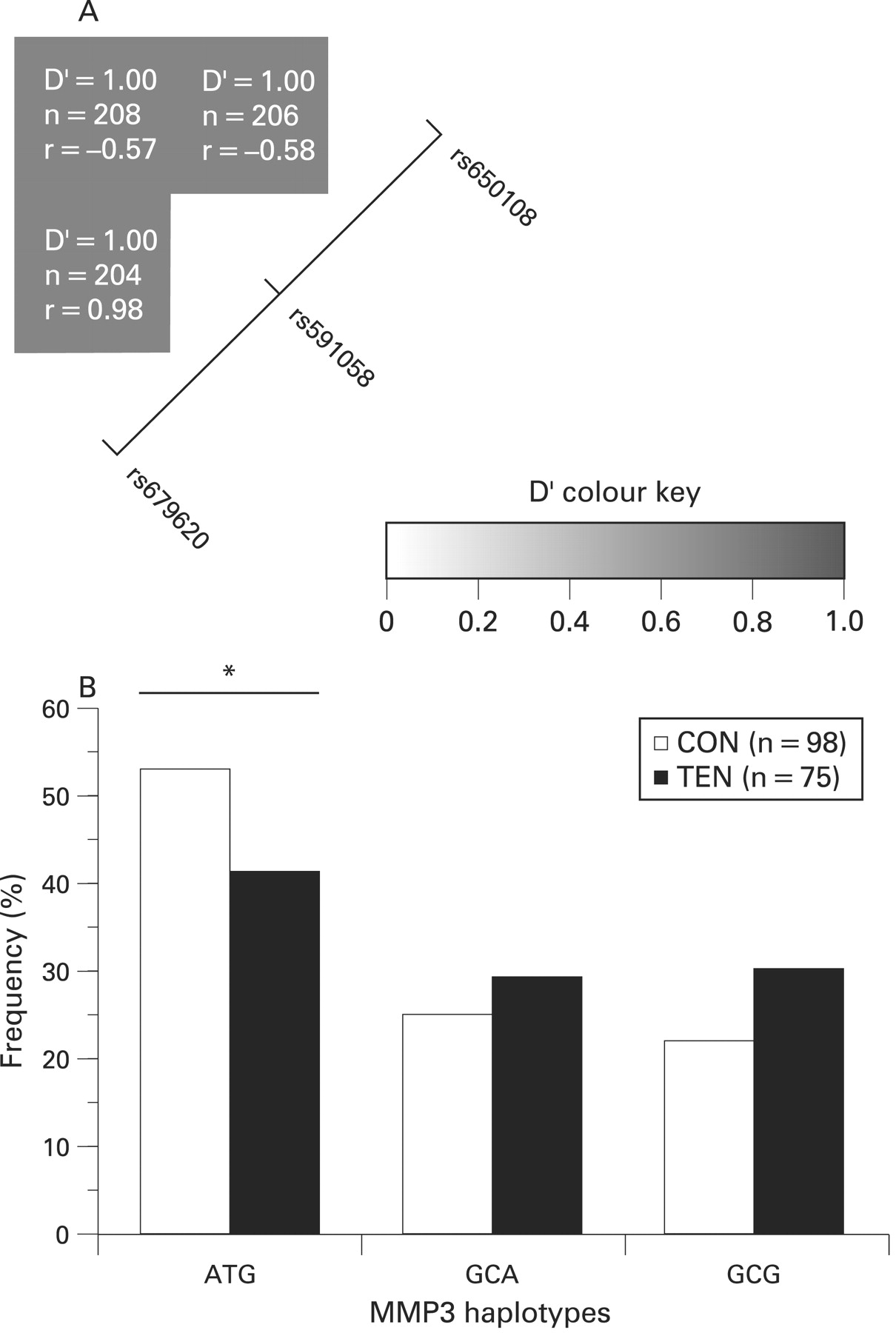
The two SNPs rs679620 (A/G) and rs591058 (T/C) are in almost perfect linkage disequilibrium (LD), allele G in the former corresponds to allele C in the latter, with only one heterozygous individual for rs679620 being TT for rs591058. The D′ measure was 1 (p<0.001) and the coefficient of correlation, r, was 0.98. Both SNPs were also in high LD with rs650108, D′ = 1 (p<0.001). The coefficient of correlation of rs650108 is −0.57 and −0.58 with rs679620 and rs591058 respectively (figure 3A). Similar values were obtained when the CON, TEN and RUP subjects were analysed separately (results not shown).
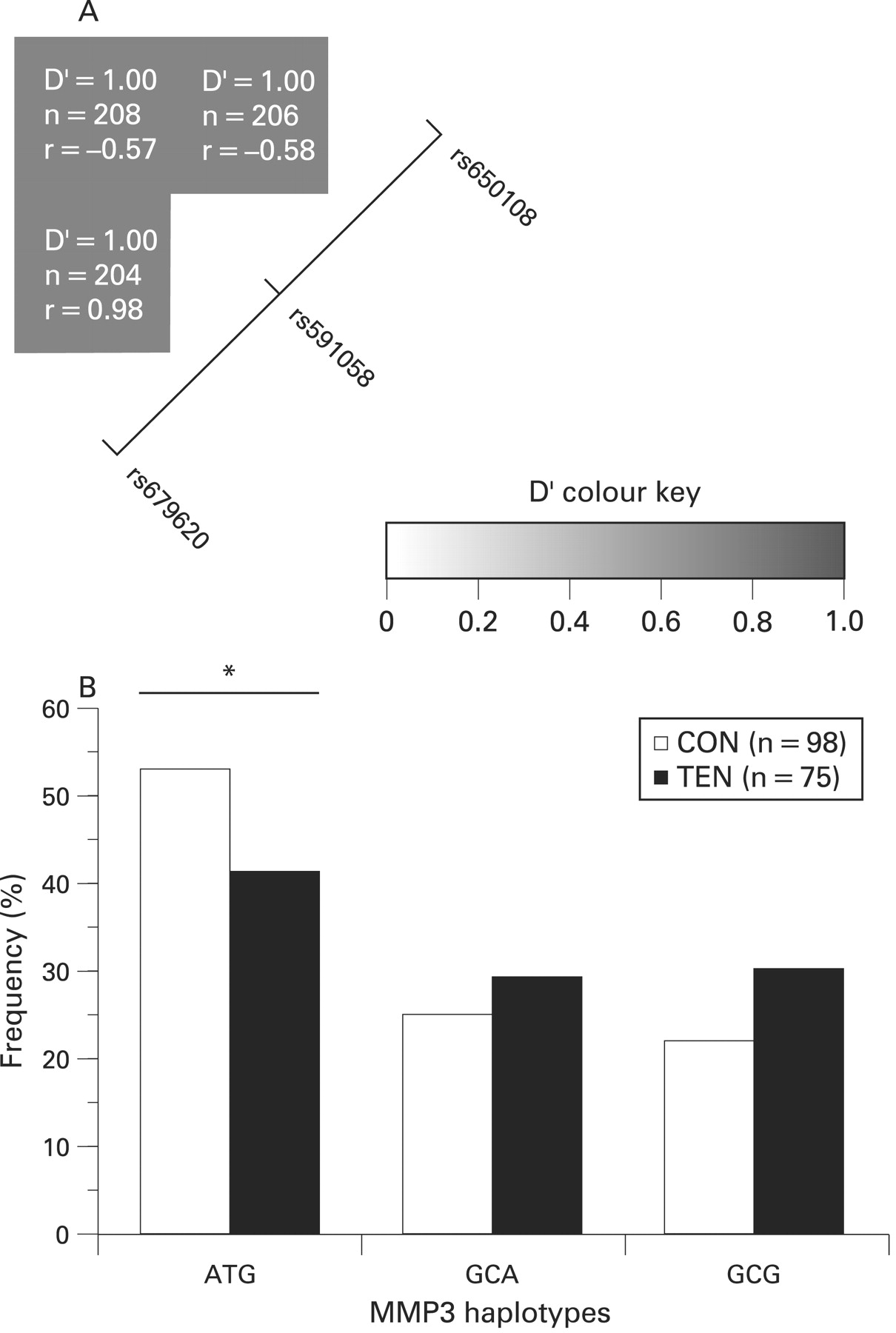
(A) Linkage disequilibrium (LD) structure of the three single nucleotide polymorphisms (SNPs), rs679620, rs591058 and rs650108, within the MMP3 gene. The LD map was constructed using the combined genotype data from the two Achilles tendon pathology and control groups. As indicated in colour key, the strength of LD between any two SNPs is indicated by the colour of the cells. The D′ values, sample sizes (n) and the coefficient of correlations (r) between pairs of markers are given within each cell. A positive correlation (r) represents major alleles being associated with each other, while a negative correlation (r) means the major allele of the one SNP is associated with the minor allelle of the other and vice versa. (B) Inferred haplotype frequency distributions from MMP3 rs679620, rs591058 and rs650108 in the control (CON, clear bars) and AT (TEN, solid bars) groups. Global P = 0.144. The asterisk and solid line marks the significant difference (p = 0.038) between the ATG haplotype pair. The number (n) of subjects in each group is in parenthesis.
As expected only the AT and GC haplotypes were inferred with a frequency greater than 1% from SNPs rs679620 and rs591058. Since the GG and CC genotypes of these SNPs were over-represented in the AT group (figs 2A and 2B), the GC haplotype was over-represented in this group (47% CON vs 59% TEN, p = 0.031). Since both these MMP3 SNPs were also in high LD with rs650108 (fig 3A), only three of the eight possible haplotypes (ATG, GCA and GCG) containing the three SNPs were inferred with a frequency more than 1%. The ATG haplotype (53% CON vs 41% TEN) was significantly under-represented in the TEN group (p = 0.038) (fig 3B).
MMP3 and COL5A1 gene-gene interaction
September et al10 have previously shown that the CC genotype of the COL5A1 BstUI RFLP (rs12722) was under-represented in patients with chronic AT. MMP3 SNP rs679620 together with the COL5A1 SNP formed the best pair of genotypes for estimating risk for AT. The genotype pairs together with their frequencies and estimated risk are summarised in table 3. The MMP3 rs679620 A allele (AA or AG genotype) combined with the COL5A1 rs12722 CC genotype had the lowest risk for AT. All the possible A and C allele combinations were associated with the lowest risk. In support of this, the MMP3 rs679620 G/A and COL5A1 rs12722 C/T allele combinations (pseudo-haplotypes) were significantly associated with TEN and CON status (global-stat = 10.4, df = 3, p = 0.016) (fig 4). The A+C allele combination was significantly associated with the controls (30% CON vs 15% TEN, p = 0.002), while the G+T allele combination was significantly associated with TEN (25% CON vs 36% TEN, p = 0.006).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
The relative distribution of the allele combinations of MMP3 SNP rs679620 (A/G) and COL5A1 SNP rs12722 (C/T) for the control (CON, clear bars) and Achilles tendinopathy (TEN, solid bars) groups. Global stat = 10.4, df = 3 and p = 0.016. The asterisk and solid line marks the significant difference between the allele combination pairs. AC, p = 0.002 and GT, p = 0.006. The number (n) of subjects in each group is in parenthesis.
DISCUSSION
We report the first evidence that sequence variation within the MMP3 gene is associated with AT. The data also demonstrate that a significant interaction exists between the exonic MMP3 SNP (rs679620) and the COL5A1 3′-untranslated region (rs12722) polymorphism and risk of AT. Our data do not support significant association between MMP3 variants alone, or in combination with the COL5A1 variant, and Achilles tendon rupture. This suggests there are different inherent predispositions to both AT and Achilles rupture. Interestingly, a similar observation has been recently reported with a RFLP in the COL5A1 gene that is associated with AT but not with ruptures.9
The apparent dissociation in genetic influence between AT and rupture is intriguing. It is known that tissue obtained during surgery to repair an acute Achilles rupture will reveal evidence of pre-existing tendon pathology.26 In contrast it has been proposed that acute tendinopathy may occur by different molecular mechanisms associated with the ageing of “normal” tendon tissue27 28 and hence may explain a different genetic predisposition to that of acute rupture. This hypothesis would challenge the assumption that similarities in gross histological appearances have the same underlying molecular mechanism of injury although this requires further investigation and verification.
All three MMP3 polymorphisms (rs679620, rs591058 and rs650108) investigated in this study have been found to associate with AT. As single loci, the rs679620 polymorphism, GG genotype, and rs591058 polymorphism, CC genotype, co-segregate with AT with odds ratios of 2.5 and 2.3 respectively. The rs650108 SNP, AA genotype has a higher odds ratio (4.9) but only seven cases possessed this genotype due to its low allele frequency in the population. When analysed as an inferred haplotype the ATG sequence combination of these three SNPs is significantly under-represented in AT cases compared to controls and it may be that this haplotype protects against the development of AT. Genotype and allele frequencies of the three SNPs investigated in this study were similar to previously reported values from databases hosted by the NCBI and Ensembl as well as from Beyzade et al.17
Although our investigation has focused on variation within the MMP3 gene and predisposition to AT, a number of previous studies have found that the expression of MMP3 RNA is reduced in both degenerative20 and ruptured Achilles samples.29 However, why this occurs is still not fully understood but reduction in MMP3 RNA or protein, without a corresponding drop in the tissue inhibitors of metalloproteins (TIMPS), might lead to an increase in proteoglycans30 which have been documented in degenerative tendons.31 Although reduced MMP3 expression or activity might lead to tendon pathology, elevated levels of MMP3 may also underlie predisposition to tendinopathy. This idea is supported by the observation that the fluoroquinolone based antibiotic ciprofloxacin, which can induce Achilles tendon pain and rupture in some individuals,32 has also been reported to substantially increase MMP3 RNA in IL-1β treated human tendon cells in culture.33
It must be remembered, of course, that allelic association to a trait or disease does not necessarily infer cause. Accordingly, this study does not address possible mechanisms of how the MMP3 polymorphisms, alone, as a haplotype, or in the case of rs679620, in combination with the COL5A1 rs12722 SNP (BstUI RFLP), might lead to pathology. However, the rs679620 variant of MMP3 is a non-synonymous polymorphism. Specifically, a glutamate residue is coded for by possession of the G allele (GAA codon) at the rs679620 loci and a lysine residue is encoded for by the A allele (AAA codon).34 Although both residues are polar the glutamate side chain is negatively charged compared to the positive charge on lysine.35 The residue sits at position 45 from the start of the polypeptide chain34 and the first 82 residues, incorporating the propeptide,36 are cleaved by a proteinase during the processing of proMMP3 into mature MMP3.37 It might be possible that appropriate removal of the propeptide has some dependency on the presence of either a Lys or Glu at position 45 and hence influence downstream function of the mature MMP3 enzyme. Beyzade et al17 have however demonstrated using clustal alignment analysis that although most of the flanking amino acids are conserved across species, the Glu45 amino acid is not.
It is possible that this association is due to genetic linkage between the non-synonymous MMP3 polymorphism (rs679620) and other polymorphisms within the MMP3 gene and flanking sequences. In support of this, the three SNPs investigated in this study, which spanned most of the gene, were in high linkage disequilibrium (D′ = 1) with each other. In addition a SNP within exon 2 of the MMP3 gene is also tightly linked (D′>0.9) to the well characterised functional −1612 −/T (5A/6A) and −1986 A/G polymorphisms within the promoter.17 The transcriptional activity of the 5A allele is higher than the 6A allele.16 Based on our and other investigator’s linkage data within this locus,17 we speculate that the non-synonymous SNP rs679620 investigated in this study is in high linkage disequilibrium with the functional 1612 −/T (5A/6A) polymorphism.
Of the 23 MMPs in humans, nine of their genes form a cluster on the long arm of chromosome 11. The MMP3 gene is part of this cluster and due to the nature of genetic association studies we can not exclude that one of the other eight MMP genes are involved in the pathogenesis of AT. Recently Ye et al18 have demonstrated that variants within MMP3, together with three other MMP genes within this cluster, namely MMP-7, -12 and -13, are associated with rheumatoid arthritis.
Our data demonstrate that the MMP3 variants investigated in this study interact with the COL5A1 rs12722 polymorphism in modifying the risk of AT. Although AT is likely to be a complex condition involving a number of gene-gene and gene-environment interactions21 there have so far been no such reports of any gene-gene interactions that relate to increased risk of AT. At present we are unable to provide a detailed mechanistic explanation for the interaction of MMP3 variants with the rs12722 polymorphism of COL5A1 but it is interesting to note that type V collagen is a reported substrate for MMP314 and may influence its synthesis and activation.38 Hence, individuals that carry risk variants within both of these genes may disrupt possible interactions between type V collagen and MMP3 during catalysis leading to a heightened risk of AT. Furthermore, future studies might attempt to establish whether there is differential expression of the MMP3 and COL5A1 genes in tendinopathy patients carrying the G and T alleles of SNPs rs679620 and rs12722, respectively, when compared to noncarriers. This data would assist in establishing whether these genes are causally implicated in AT.
In addition to the genetic factors identified in this and other studies,8–10 non-genetic intrinsic factors, as well as extrinsic factors, have been implicated in the aetiology of Achilles tendon pathology.39 As previously discussed8 we cannot exclude the possibility that an interaction of weight and/or physical activity exposure with the susceptibility genotype was responsible for the development of Achilles tendon pathology symptoms.
A limitation of this study was that the symptomatic subjects were significantly heavier than the control subjects. Since this was a retrospective study, the body weight of the subjects at the time of injury is unknown. Anecdotally, many subjects reported increases in their body weight after injury as a result of a decrease in physical activity. An additional limitation of this study was the modest sample size.
In conclusion, we report the first evidence that variation within the human MMP3 gene is associated with AT. We have also established, for the first time, that the MMP3 variants we have studied interact with a variant within the COL5A1 gene to increase risk of AT. We suggest repetition of this work in other, larger population studies to confirm our associations.
What is already known about this topic
Both TNC and COL5A1 gene variants have been associated with an increased risk of Achilles tendinopathy.
What this study adds
Our data demonstrate, for the first time, that variants within the matrix metalloprotein 3 gene (MMP3) can significantly modify the risk of tendinopathy. We also show that a variant within the COL5A1 gene interacts with MMP3 variants to modify risk.
Acknowledgments
The authors would like to thank Dr GG Mokone for assistance in the recruitment of the subjects and the phenotypic data collection for the subjects.
REFERENCES
Footnotes
Funding: This study was supported in part by funds from the National Research Foundation (NRF) of South Africa (grants no: FA2005021700015 and FA2007032700010), University of Cape Town, University of Northampton and the South African Medical Research Council (MRC).
Competing interests: None.
Ethics approval: This study was approved by the Research Ethics Committees of the Faculty of Health Sciences within the University of Cape Town, South Africa and the University of Northampton, UK.